

AMPK activation protects against prostate cancer by inducing a catabolic cellular state
- PMID: 37061917
- PMCID: PMC10576838
- DOI: 10.1016/j.celrep.2023.112396
AMPK activation protects against prostate cancer by inducing a catabolic cellular state
Abstract
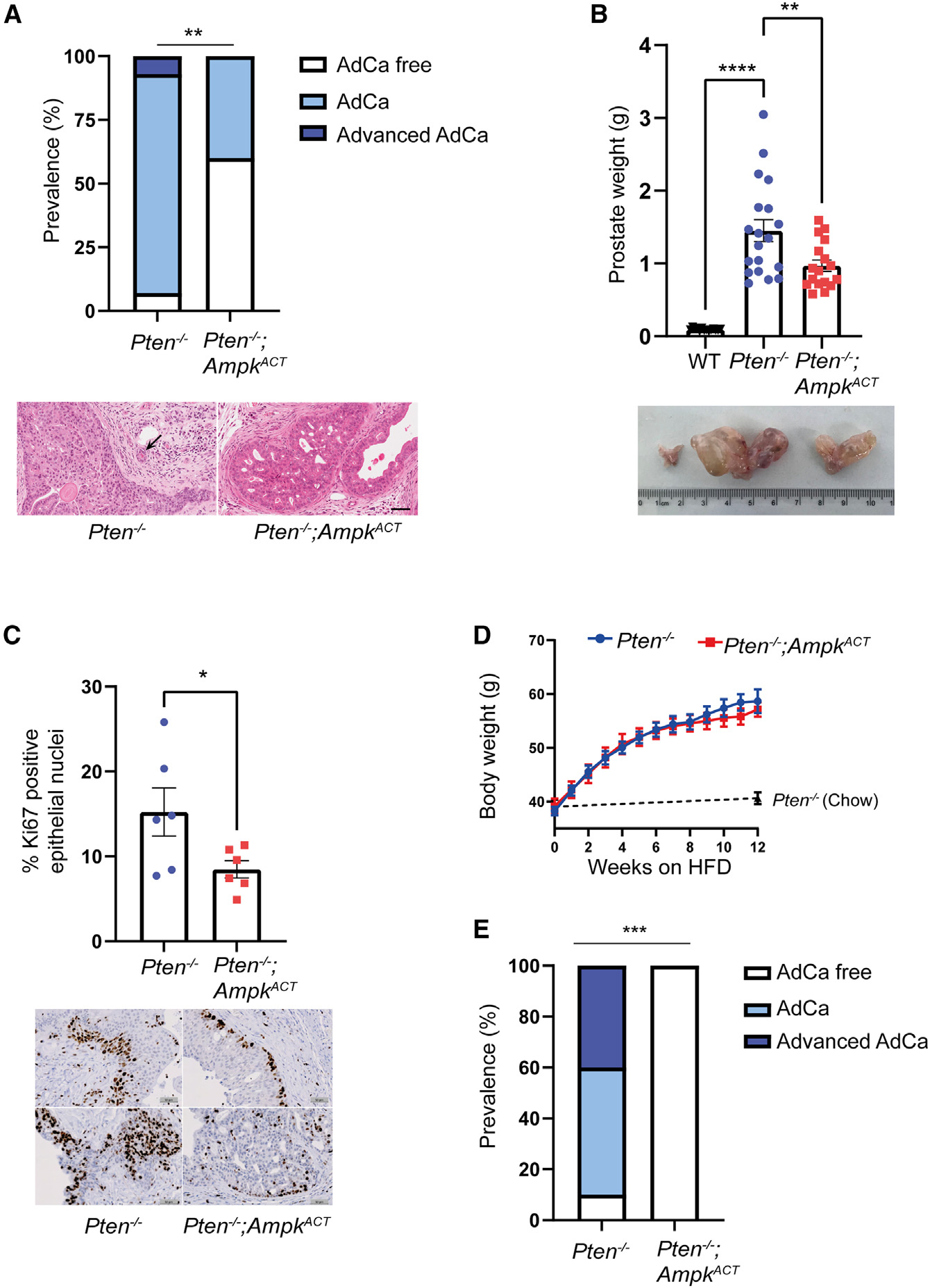
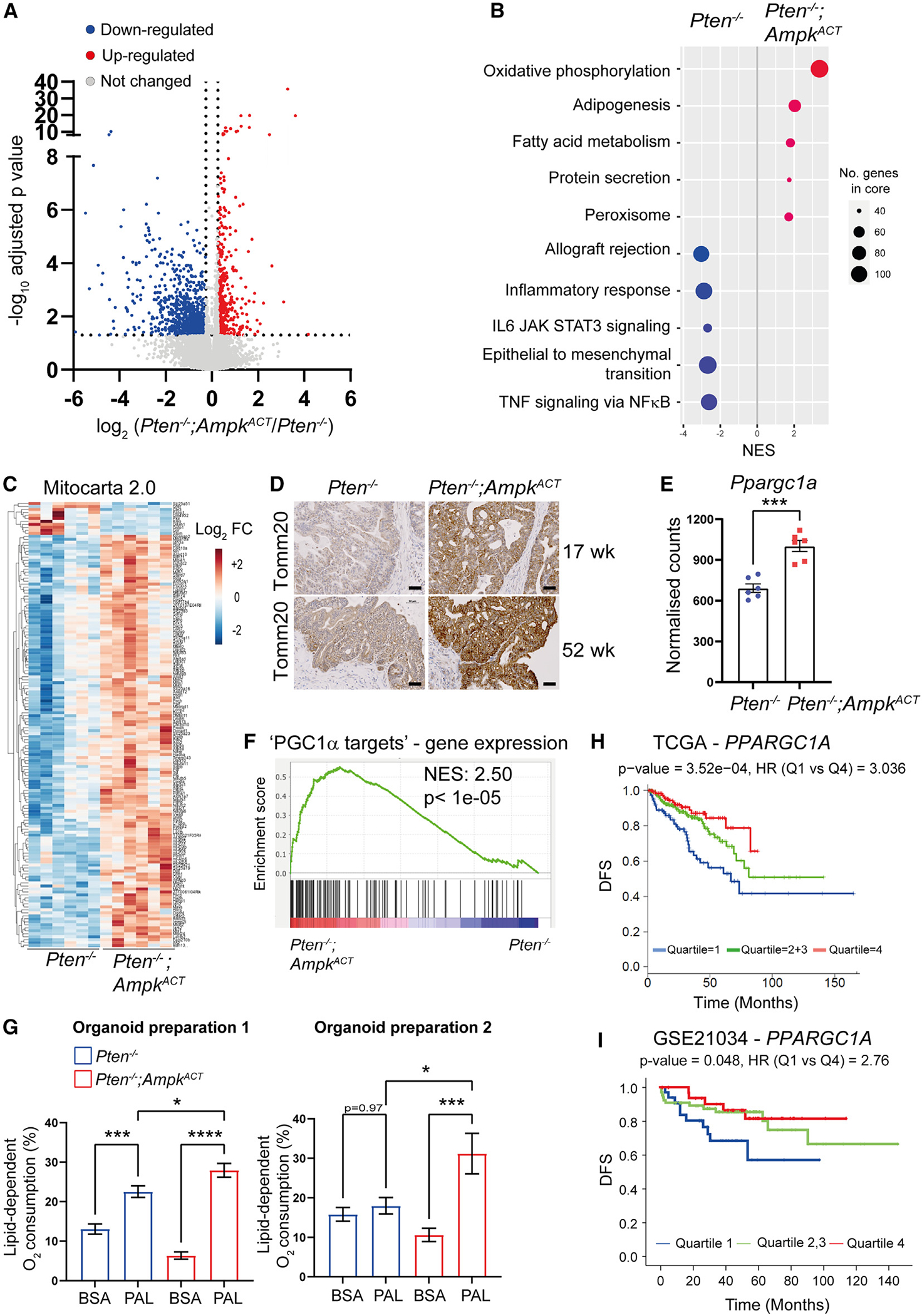
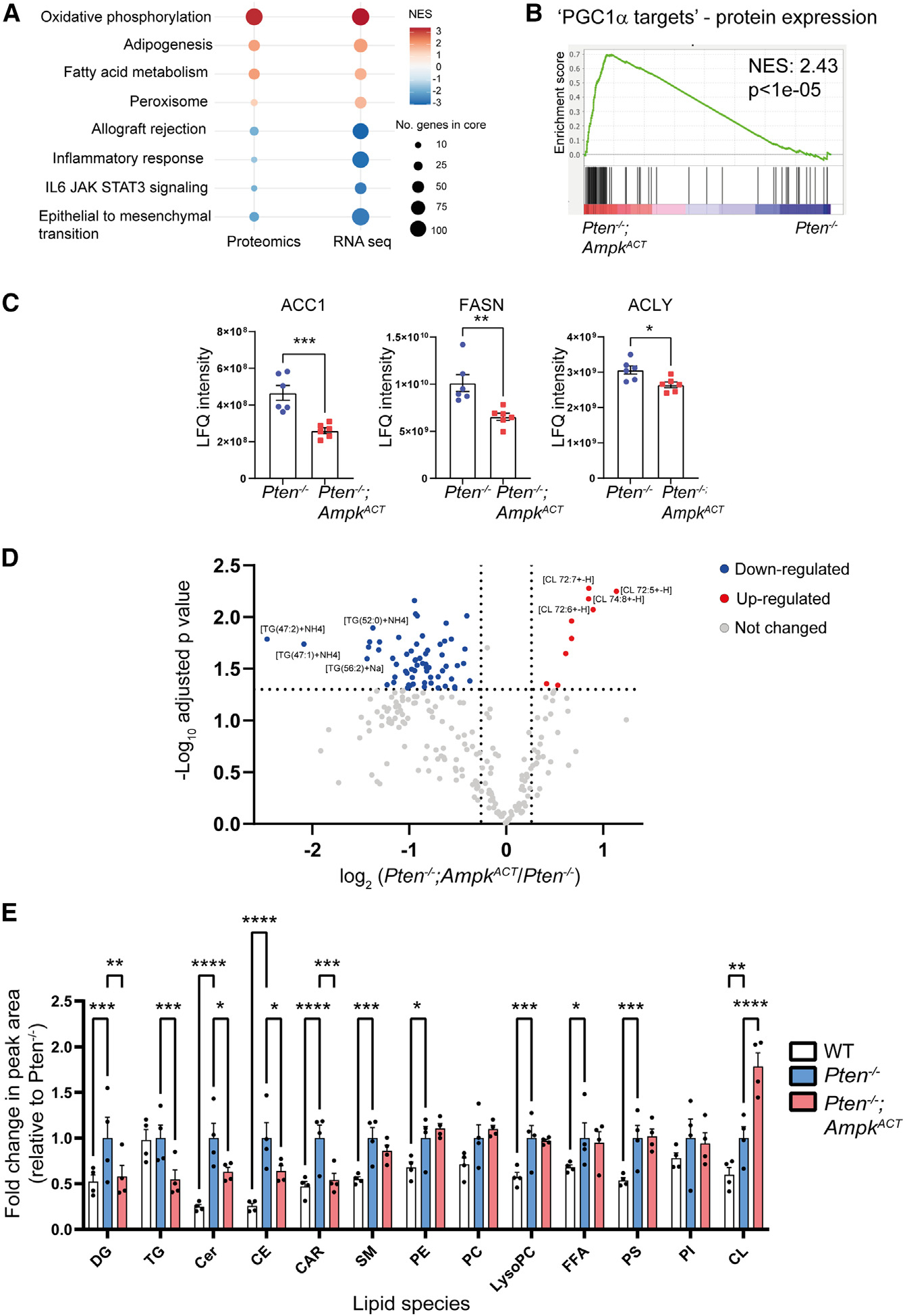
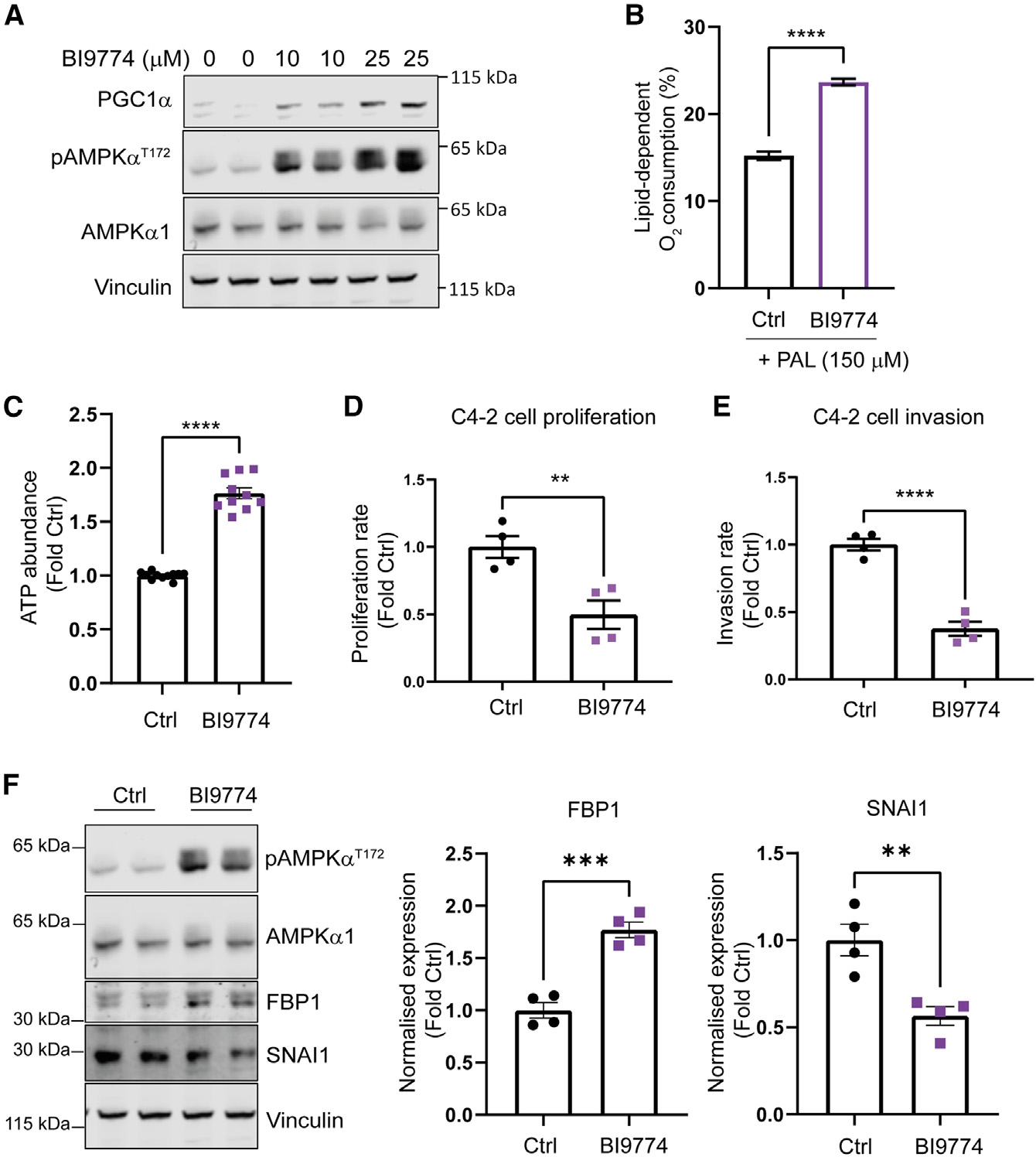
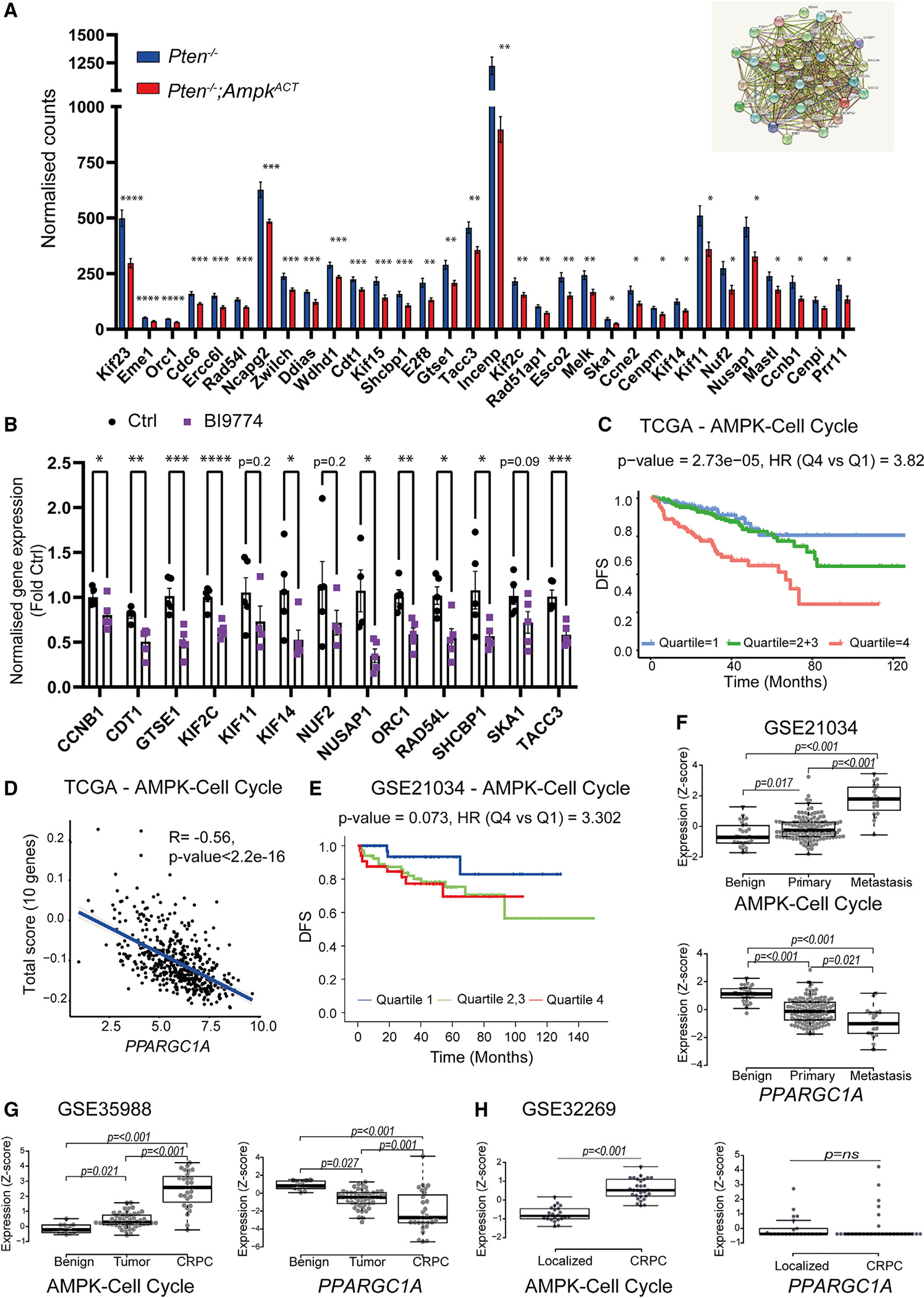
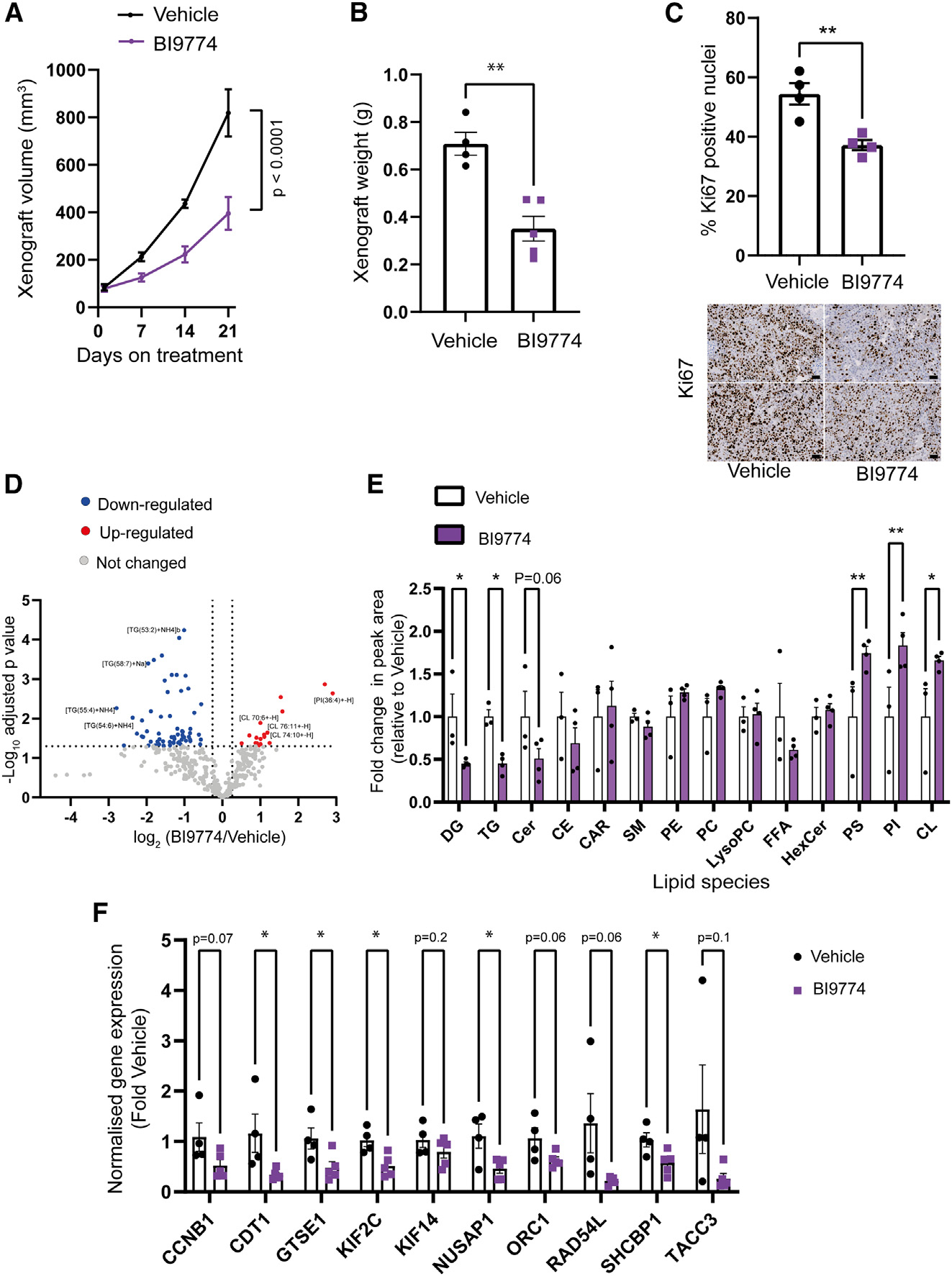
Emerging evidence indicates that metabolic dysregulation drives prostate cancer (PCa) progression and metastasis. AMP-activated protein kinase (AMPK) is a master regulator of metabolism, although its role in PCa remains unclear. Here, we show that genetic and pharmacological activation of AMPK provides a protective effect on PCa progression in vivo. We show that AMPK activation induces PGC1α expression, leading to catabolic metabolic reprogramming of PCa cells. This catabolic state is characterized by increased mitochondrial gene expression, increased fatty acid oxidation, decreased lipogenic potential, decreased cell proliferation, and decreased cell invasiveness. Together, these changes inhibit PCa disease progression. Additionally, we identify a gene network involved in cell cycle regulation that is inhibited by AMPK activation. Strikingly, we show a correlation between this gene network and PGC1α gene expression in human PCa. Taken together, our findings support the use of AMPK activators for clinical treatment of PCa to improve patient outcome.
Keywords: AMPK; CP: Cancer; CP: Metabolism; PGC1α; cell cycle regulation; fatty acid oxidation; high-fat diet; lipogenesis; metabolism; metastasis; mitochondria; prostate cancer.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- MC_U120027537/MRC_/Medical Research Council/United Kingdom
- R01 CA197160/CA/NCI NIH HHS/United States
- MC-A654-5QB10/MRC_/Medical Research Council/United Kingdom
- BB/W009633/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MC_UP_1605/5/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
