

Peptide chain release factor DIG8 regulates plant growth by affecting ROS-mediated sugar transportation in Arabidopsis thaliana
- PMID: 37063204
- PMCID: PMC10102589
- DOI: 10.3389/fpls.2023.1172275
Peptide chain release factor DIG8 regulates plant growth by affecting ROS-mediated sugar transportation in Arabidopsis thaliana
Abstract
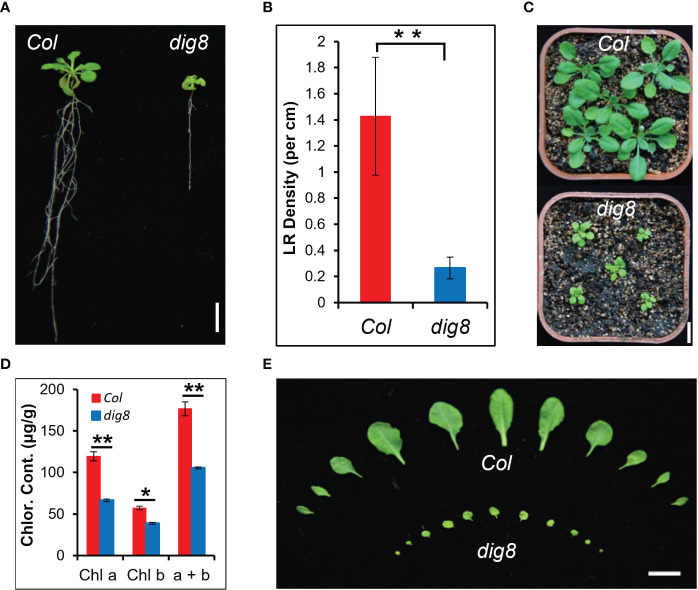
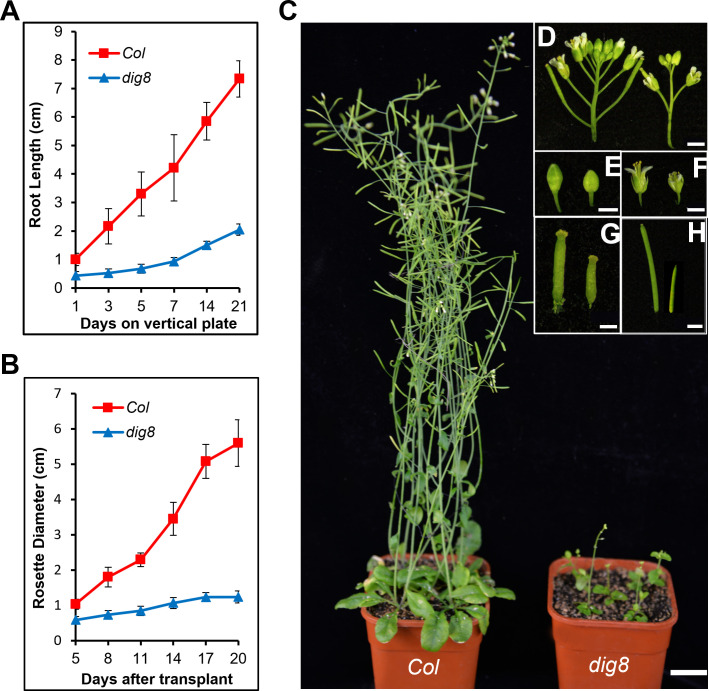
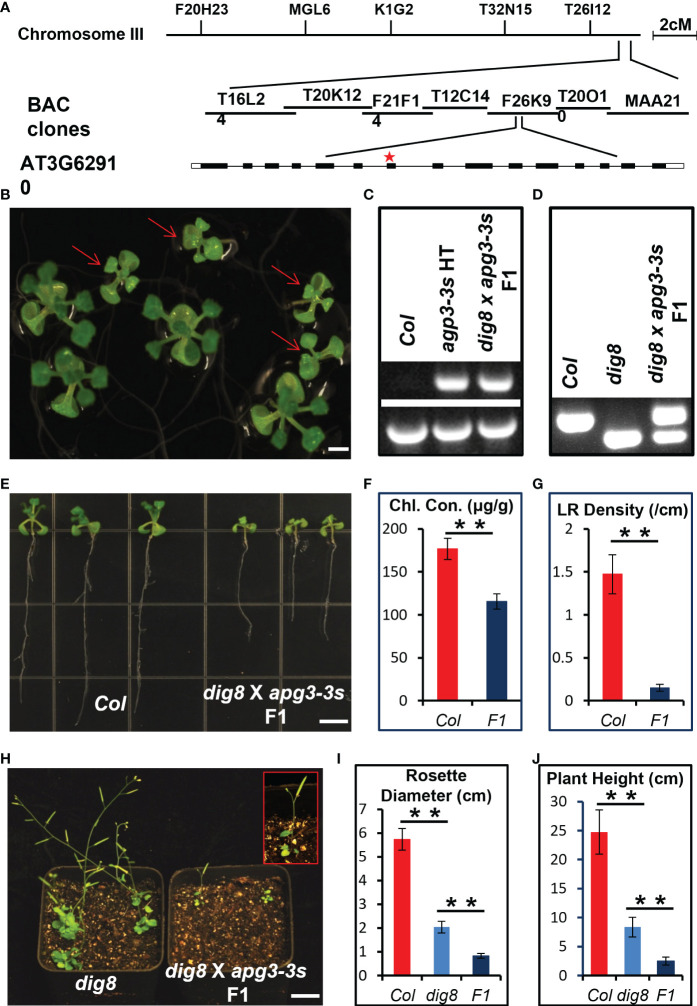
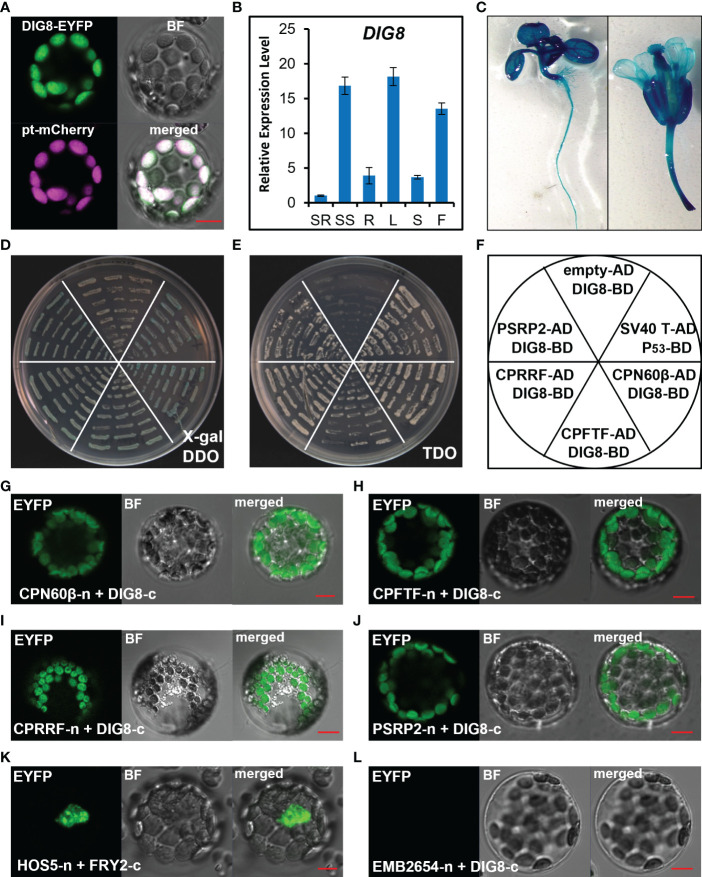
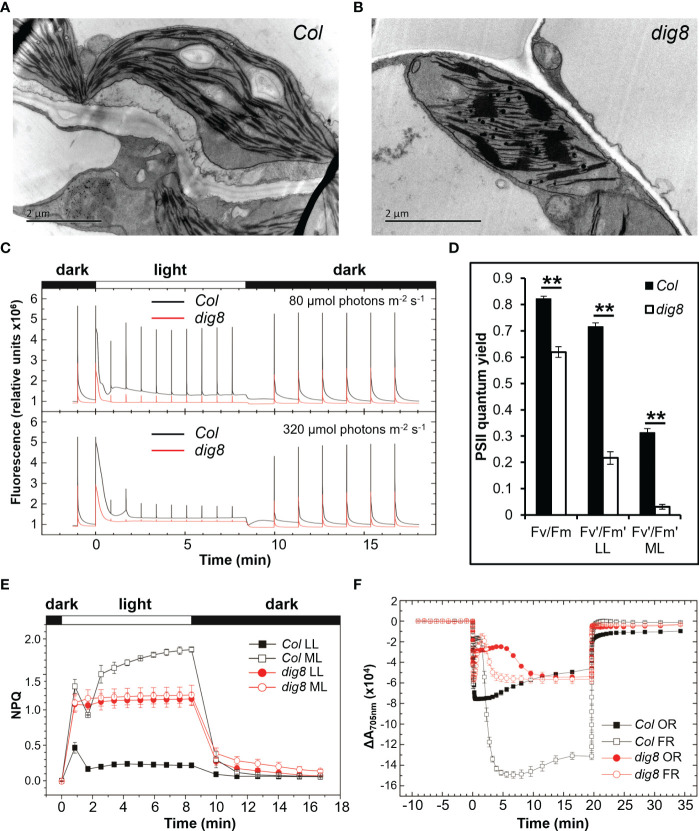
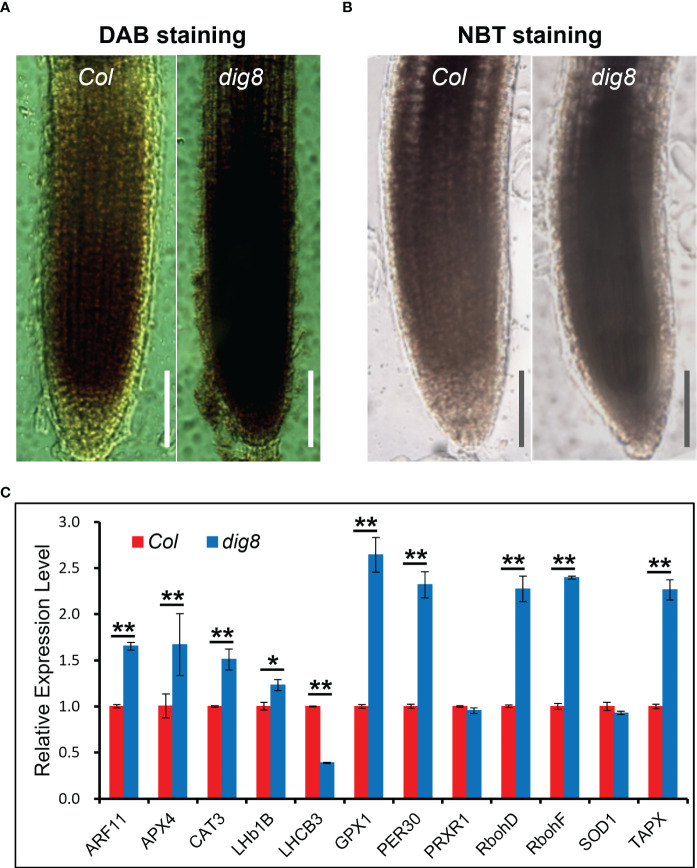
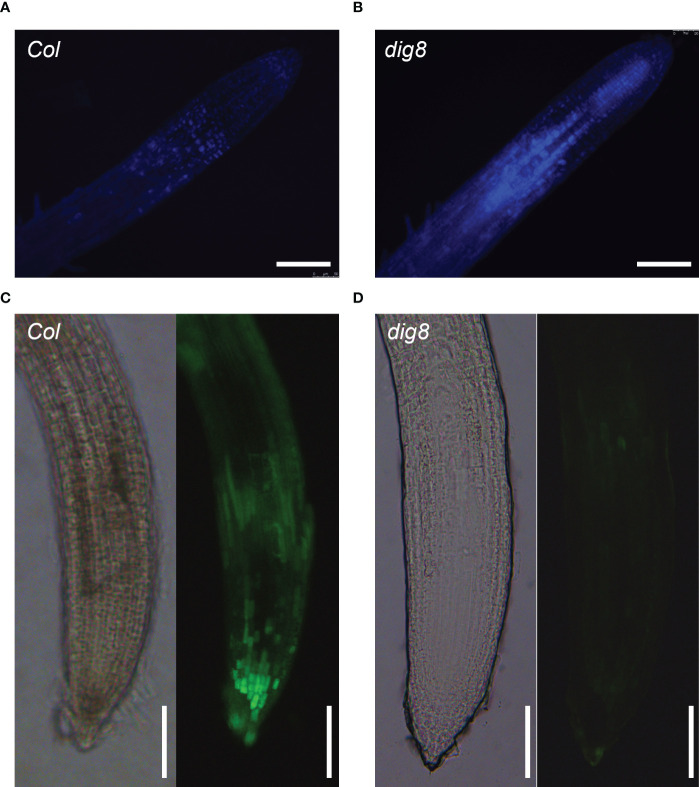
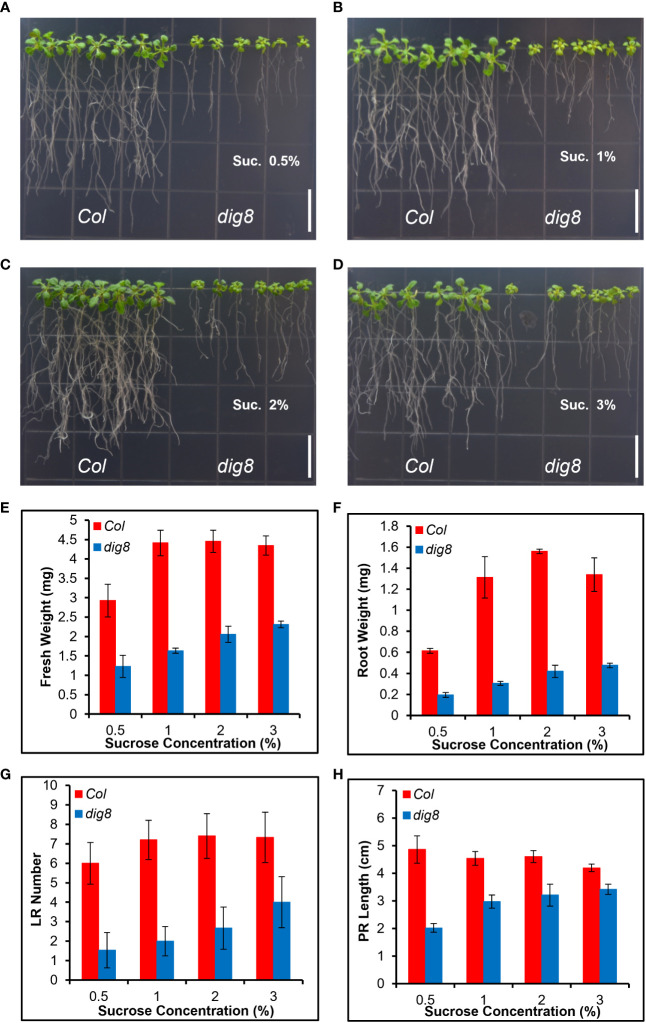
Chloroplasts have important roles in photosynthesis, stress sensing and retrograde signaling. However, the relationship between chloroplast peptide chain release factor and ROS-mediated plant growth is still unclear. In the present study, we obtained a loss-of-function mutant dig8 by EMS mutation. The dig8 mutant has few lateral roots and a pale green leaf phenotype. By map-based cloning, the DIG8 gene was located on AT3G62910, with a point mutation leading to amino acid substitution in functional release factor domain. Using yeast-two-hybrid and BiFC, we confirmed DIG8 protein was characterized locating in chloroplast by co-localization with plastid marker and interacting with ribosome-related proteins. Through observing by transmission electron microscopy, quantifying ROS content and measuring the transport efficiency of plasmodesmata in dig8 mutant, we found that abnormal thylakoid stack formation and chloroplast dysfunction in the dig8 mutant caused increased ROS activity leading to callose deposition and lower PD permeability. A local sugar supplement partially alleviated the growth retardation phenotype of the mutant. These findings shed light on chloroplast peptide chain release factor-affected plant growth by ROS stress.
Keywords: ROS; callose; chloroplast; peptide chain release factor; sugar transportation.
Copyright © 2023 Zhang, Han, Han, Zhang, Xiong and Chen.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Characterization of the snowy cotyledon 1 mutant of Arabidopsis thaliana: the impact of chloroplast elongation factor G on chloroplast development and plant vitality.Plant Mol Biol. 2006 Mar;60(4):507-18. doi: 10.1007/s11103-005-4921-0. Plant Mol Biol. 2006. PMID: 16525888
-
Chloroplast ribosome release factor 1 (AtcpRF1) is essential for chloroplast development.Plant Mol Biol. 2007 Jul;64(5):481-97. doi: 10.1007/s11103-007-9166-7. Epub 2007 Apr 21. Plant Mol Biol. 2007. PMID: 17450416
-
A Putative Chloroplast Thylakoid Metalloprotease VIRESCENT3 Regulates Chloroplast Development in Arabidopsis thaliana.J Biol Chem. 2016 Feb 12;291(7):3319-32. doi: 10.1074/jbc.M115.681601. Epub 2015 Dec 23. J Biol Chem. 2016. PMID: 26702056 Free PMC article.
-
The Plastid Terminal Oxidase is a Key Factor Balancing the Redox State of Thylakoid Membrane.Enzymes. 2016;40:143-171. doi: 10.1016/bs.enz.2016.09.002. Epub 2016 Oct 12. Enzymes. 2016. PMID: 27776780 Review.
-
Chloroplast ROS and stress signaling.Plant Commun. 2021 Nov 9;3(1):100264. doi: 10.1016/j.xplc.2021.100264. eCollection 2022 Jan 10. Plant Commun. 2021. PMID: 35059631 Free PMC article. Review.
Cited by
-
A CRISPR/Cas9-induced male-sterile line facilitating easy hybrid production in polyploid rapeseed (Brassica napus).Hortic Res. 2024 May 28;11(7):uhae139. doi: 10.1093/hr/uhae139. eCollection 2024 Jul. Hortic Res. 2024. PMID: 38988621 Free PMC article.
References
LinkOut - more resources
Full Text Sources