

Infectious viruses from transfected SARS-CoV-2 genomic RNA
- PMID: 37064222
- PMCID: PMC10098207
- DOI: 10.3389/fbioe.2023.1129111
Infectious viruses from transfected SARS-CoV-2 genomic RNA
Abstract
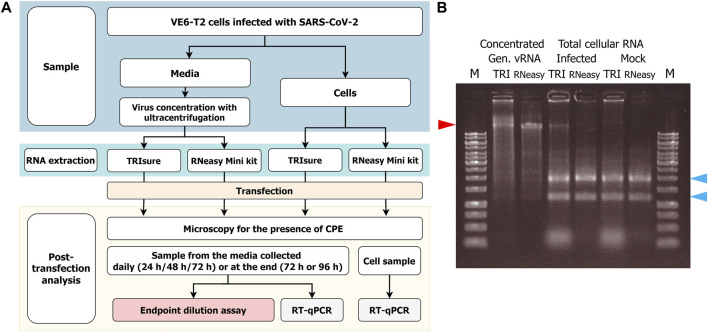
SARS-CoV-2 emerged at the end of 2019, and like other novel pathogens causing severe symptoms, WHO recommended heightened biosafety measures for laboratories working with the virus. The positive-stranded genomic RNA of coronaviruses has been known to be infectious since the 1970s, and overall, all experiments with the possibility of SARS-CoV-2 propagation are carried out in higher containment level laboratories. However, as SARS-CoV-2 RNA has been routinely handled in BSL-2 laboratories, the question of the true nature of RNA infectiousness has risen along with discussion of appropriate biosafety measures. Here, we studied the ability of native SARS-CoV-2 genomic RNA to produce infectious viruses when transfected into permissive cells and discussed the biosafety control measures related to these assays. In transfection assays large quantities of genomic vRNA of SARS-CoV-2 was required for a successful production of infectious viruses. However, the quantity of vRNA alone was not the only factor, and especially when the transfected RNA was derived from infected cells, even small amounts of genomic vRNA was enough for an infection. Virus replication was found to start rapidly after transfection, and infectious viruses were detected in the cell culture media at 24 h post-transfection. In addition, silica membrane-based kits were shown to be as good as traditional TRI-reagent based methods in extracting high-quality, 30 kb-long genomic vRNA. Taken together, our data indicates that all transfection experiments with samples containing genomic SARS-CoV-2 RNA should be categorized as a propagative work and the work should be conducted only in a higher containment BSL-3 laboratory.
Keywords: SARS-CoV-2; biosafety; genomic RNA; infectivity; transfection.
Copyright © 2023 Väisänen, Jiang, Laine, Waris, Julkunen and Österlund.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
A Universal Next-Generation Sequencing Protocol To Generate Noninfectious Barcoded cDNA Libraries from High-Containment RNA Viruses.mSystems. 2016 Jun 7;1(3):e00039-15. doi: 10.1128/mSystems.00039-15. eCollection 2016 May-Jun. mSystems. 2016. PMID: 27822536 Free PMC article.
-
No Evidence of Infectious SARS-CoV-2 in Human Milk: Analysis of a Cohort of 110 Lactating Women.medRxiv [Preprint]. 2021 Apr 7:2021.04.05.21254897. doi: 10.1101/2021.04.05.21254897. medRxiv. 2021. Update in: Pediatr Res. 2022 Oct;92(4):1140-1145. doi: 10.1038/s41390-021-01902-y. PMID: 33851178 Free PMC article. Updated. Preprint.
-
Biorisk Management for SARS-CoV-2 Research in a Biosafety Level-3 Core Facility.Methods Mol Biol. 2022;2452:441-464. doi: 10.1007/978-1-0716-2111-0_24. Methods Mol Biol. 2022. PMID: 35554920
-
The double-membrane vesicle (DMV): a virus-induced organelle dedicated to the replication of SARS-CoV-2 and other positive-sense single-stranded RNA viruses.Cell Mol Life Sci. 2022 Jul 16;79(8):425. doi: 10.1007/s00018-022-04469-x. Cell Mol Life Sci. 2022. PMID: 35841484 Free PMC article. Review.
-
Laboratory containment of SARS virus.Ann Acad Med Singap. 2006 May;35(5):354-60. Ann Acad Med Singap. 2006. PMID: 16830004 Review.
References
-
- Campillo-Balderas J. A., Lazcano A., Becerra A. (2015). Viral genome size distribution does not correlate with the antiquity of the host lineages. Front. Ecol. Evol. 3, 143. 10.3389/fevo.2015.00143 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous