

Thioredoxin VdTrx1, an unconventional secreted protein, is a virulence factor in Verticillium dahliae
- PMID: 37065139
- PMCID: PMC10102666
- DOI: 10.3389/fmicb.2023.1130468
Thioredoxin VdTrx1, an unconventional secreted protein, is a virulence factor in Verticillium dahliae
Abstract
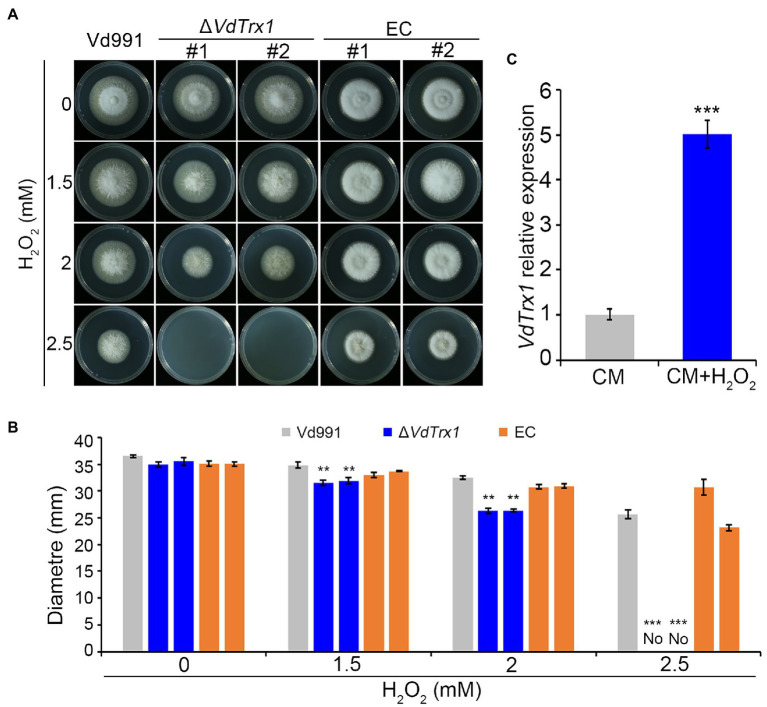
Understanding how plant pathogenic fungi adapt to their hosts is of critical importance to securing optimal crop productivity. In response to pathogenic attack, plants produce reactive oxygen species (ROS) as part of a multipronged defense response. Pathogens, in turn, have evolved ROS scavenging mechanisms to undermine host defense. Thioredoxins (Trx) are highly conserved oxidoreductase enzymes with a dithiol-disulfide active site, and function as antioxidants to protect cells against free radicals, such as ROS. However, the roles of thioredoxins in Verticillium dahliae, an important vascular pathogen, are not clear. Through proteomics analyses, we identified a putative thioredoxin (VdTrx1) lacking a signal peptide. VdTrx1 was present in the exoproteome of V. dahliae cultured in the presence of host tissues, a finding that suggested that it plays a role in host-pathogen interactions. We constructed a VdTrx1 deletion mutant ΔVdTrx1 that exhibited significantly higher sensitivity to ROS stress, H2O2, and tert-butyl hydroperoxide (t-BOOH). In vivo assays by live-cell imaging and in vitro assays by western blotting revealed that while VdTrx1 lacking the signal peptide can be localized within V. dahliae cells, VdTrx1 can also be secreted unconventionally depending on VdVps36, a member of the ESCRT-II protein complex. The ΔVdTrx1 strain was unable to scavenge host-generated extracellular ROS fully during host invasion. Deletion of VdTrx1 resulted in higher intracellular ROS levels of V. dahliae mycelium, displayed impaired conidial production, and showed significantly reduced virulence on Gossypium hirsutum, and model plants, Arabidopsis thaliana and Nicotiana benthamiana. Thus, we conclude that VdTrx1 acts as a virulence factor in V. dahliae.
Keywords: ROS scavenging; Verticillium dahliae; thioredoxin; unconventional secreted protein; virulence factor.
Copyright © 2023 Tian, Zhuang, Li, Zhu, Klosterman, Dai, Chen, Subbarao and Zhang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







Similar articles
-
Cu/Zn superoxide dismutase (VdSOD1) mediates reactive oxygen species detoxification and modulates virulence in Verticillium dahliae.Mol Plant Pathol. 2021 Sep;22(9):1092-1108. doi: 10.1111/mpp.13099. Epub 2021 Jul 9. Mol Plant Pathol. 2021. PMID: 34245085 Free PMC article.
-
VdGAL4 Modulates Microsclerotium Formation, Conidial Morphology, and Germination To Promote Virulence in Verticillium dahliae.Microbiol Spectr. 2023 Feb 14;11(1):e0351522. doi: 10.1128/spectrum.03515-22. Epub 2022 Dec 7. Microbiol Spectr. 2023. PMID: 36475739 Free PMC article.
-
The Thioredoxin GbNRX1 Plays a Crucial Role in Homeostasis of Apoplastic Reactive Oxygen Species in Response to Verticillium dahliae Infection in Cotton.Plant Physiol. 2016 Apr;170(4):2392-406. doi: 10.1104/pp.15.01930. Epub 2016 Feb 11. Plant Physiol. 2016. PMID: 26869704 Free PMC article.
-
An Overview of the Molecular Genetics of Plant Resistance to the Verticillium Wilt Pathogen Verticillium dahliae.Int J Mol Sci. 2020 Feb 7;21(3):1120. doi: 10.3390/ijms21031120. Int J Mol Sci. 2020. PMID: 32046212 Free PMC article. Review.
-
Physiological and molecular mechanism of defense in cotton against Verticillium dahliae.Plant Physiol Biochem. 2018 Apr;125:193-204. doi: 10.1016/j.plaphy.2018.02.011. Epub 2018 Feb 13. Plant Physiol Biochem. 2018. PMID: 29462745 Review.
Cited by
-
In silico analysis of secreted effectorome of the rubber tree pathogen Rigidoporus microporus highlights its potential virulence proteins.Front Microbiol. 2024 Sep 16;15:1439454. doi: 10.3389/fmicb.2024.1439454. eCollection 2024. Front Microbiol. 2024. PMID: 39360316 Free PMC article.
-
CAP superfamily proteins (VdPRYs) manipulate plant immunity and contribute to the virulence of Verticillium dahliae.Mycology. 2024 Nov 8;16(2):876-890. doi: 10.1080/21501203.2024.2419882. eCollection 2025. Mycology. 2024. PMID: 40415913 Free PMC article.
-
The acyl-CoA-binding protein VdAcb1 is essential for carbon starvation response and contributes to virulence in Verticillium dahliae.aBIOTECH. 2024 Jul 13;5(4):431-448. doi: 10.1007/s42994-024-00175-3. eCollection 2024 Dec. aBIOTECH. 2024. PMID: 39650135 Free PMC article.
-
Genomic insights into Verticillium: a review of progress in the genomics era.Front Microbiol. 2024 Oct 11;15:1463779. doi: 10.3389/fmicb.2024.1463779. eCollection 2024. Front Microbiol. 2024. PMID: 39464398 Free PMC article. Review.
-
Mitochondrial Complex Member VdNuo1 Recruits Superoxide Dismutases VdSOD2/4 to Maintain Superoxide Anion Homeostasis During Pathogenesis in Verticillium dahliae.Mol Plant Pathol. 2025 Sep;26(9):e70149. doi: 10.1111/mpp.70149. Mol Plant Pathol. 2025. PMID: 40913259 Free PMC article.
References
-
- Bui T. T., Harting R., Braus-Stromeyer S. A., Tran V. T., Leonard M., Höfer A., et al. . (2019). Verticillium dahliae transcription factors Som1 and Vta3 control microsclerotia formation and sequential steps of plant root penetration and colonisation to induce disease. New Phytol. 221, 2138–2159. doi: 10.1111/nph.15514 - DOI - PubMed