

Probing the conserved roles of cut in the development and function of optically different insect compound eyes
- PMID: 37065850
- PMCID: PMC10102356
- DOI: 10.3389/fcell.2023.1104620
Probing the conserved roles of cut in the development and function of optically different insect compound eyes
Abstract
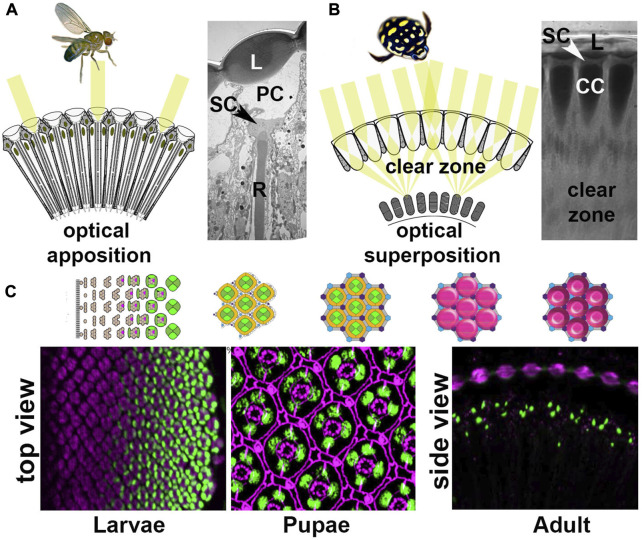
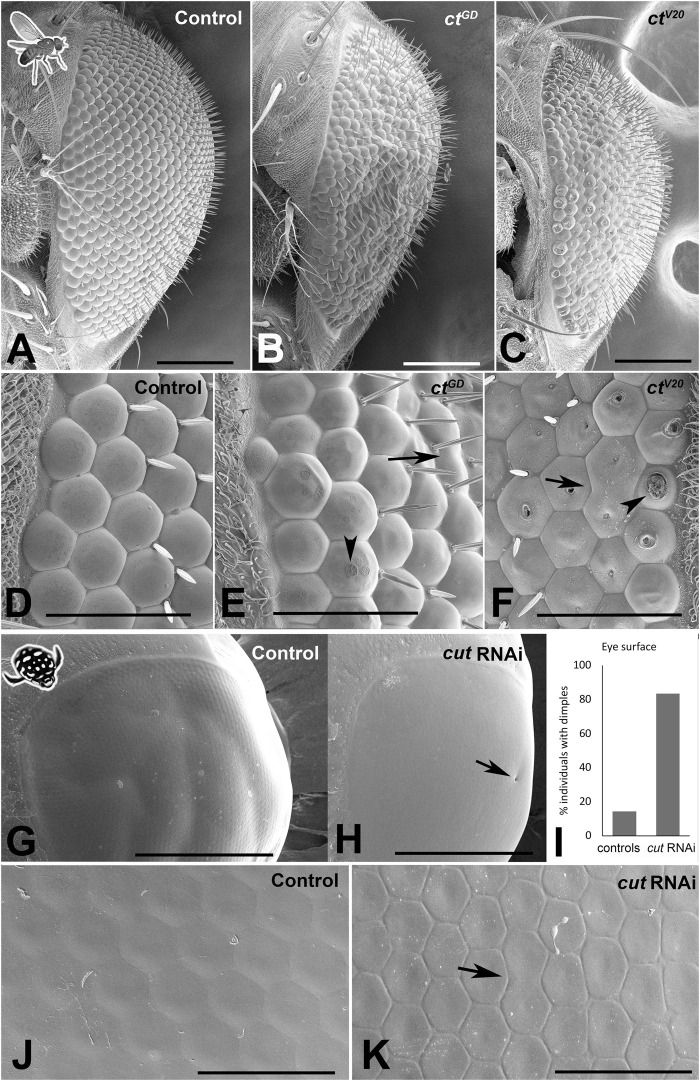
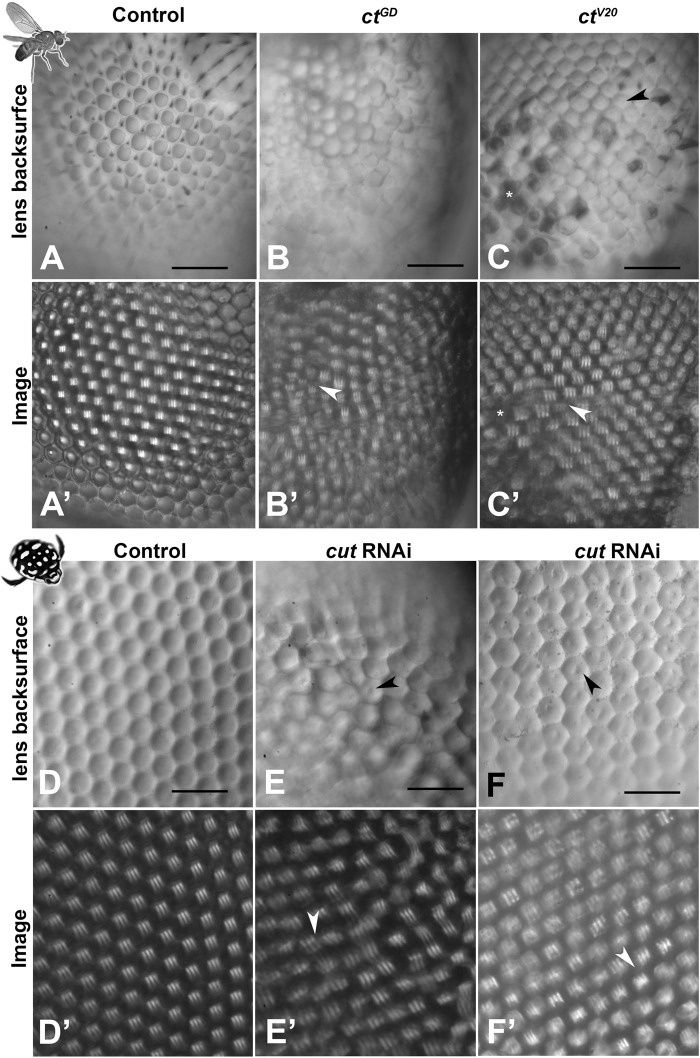
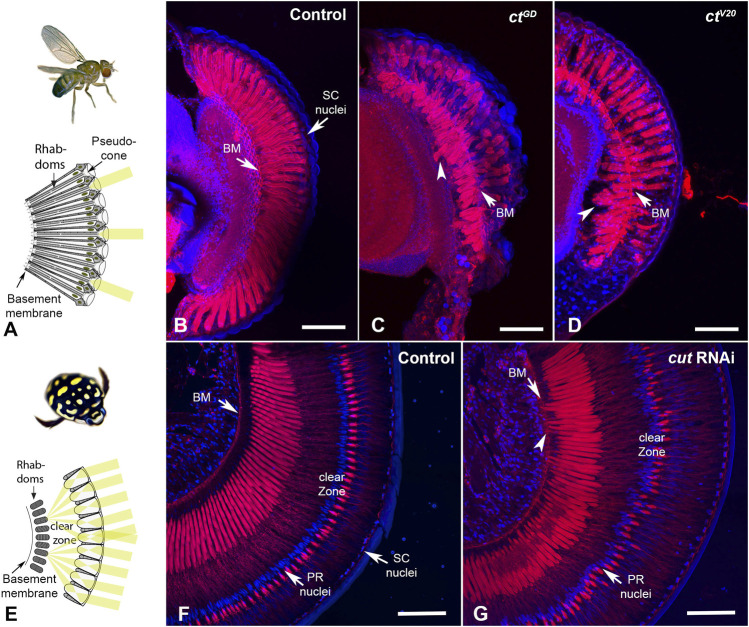
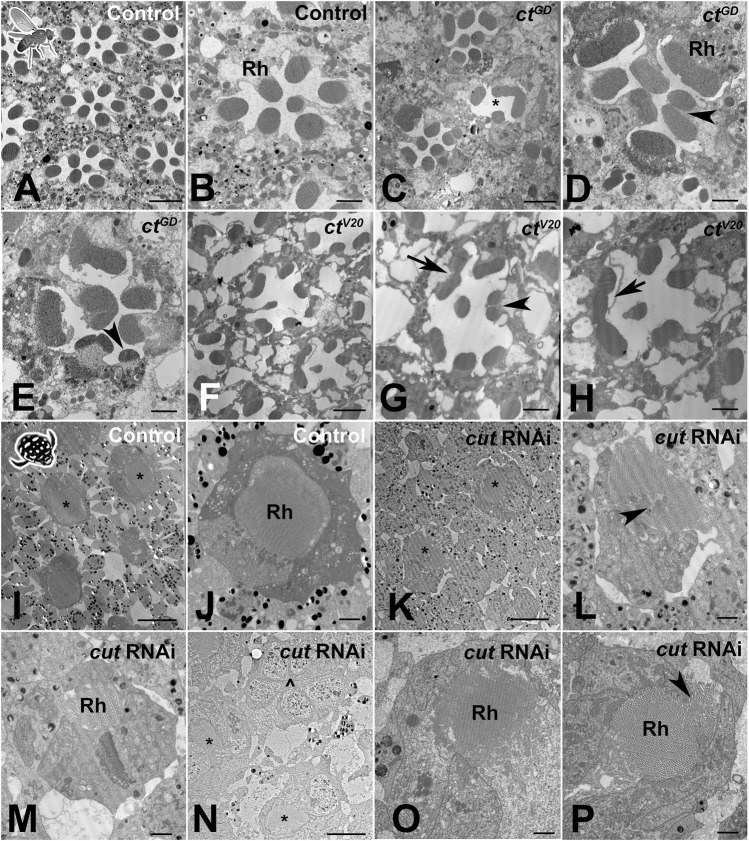
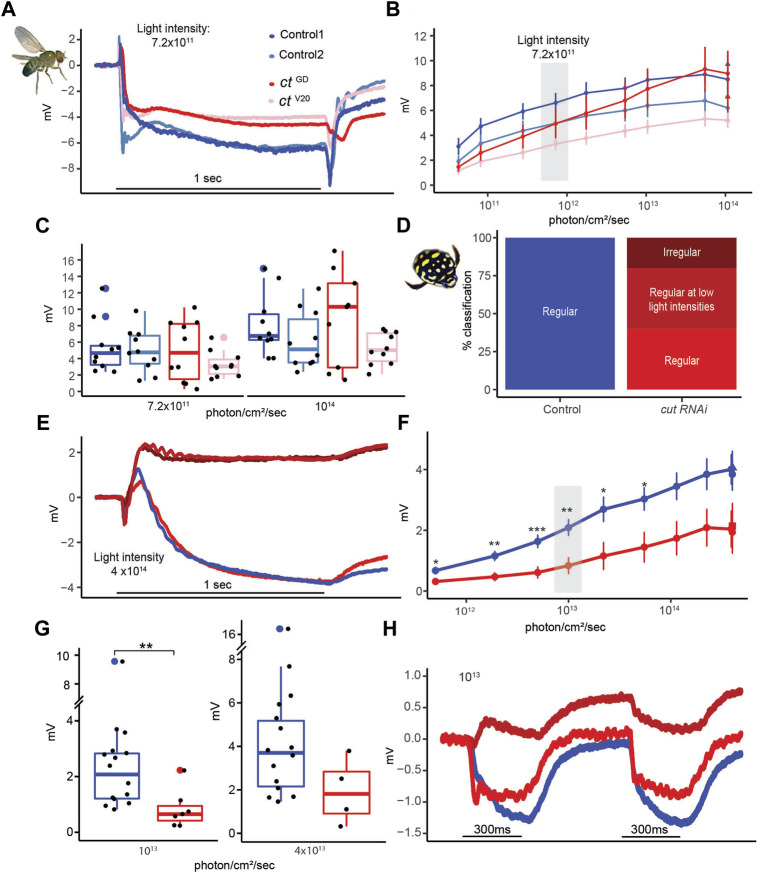
Astonishing functional diversity exists among arthropod eyes, yet eye development relies on deeply conserved genes. This phenomenon is best understood for early events, whereas fewer investigations have focused on the influence of later transcriptional regulators on diverse eye organizations and the contribution of critical support cells, such as Semper cells (SCs). As SCs in Drosophila melanogaster secrete the lens and function as glia, they are critical components of ommatidia. Here, we perform RNAi-based knockdowns of the transcription factor cut (CUX in vertebrates), a marker of SCs, the function of which has remained untested in these cell types. To probe for the conserved roles of cut, we investigate two optically different compound eyes: the apposition optics of D. melanogaster and the superposition optics of the diving beetle Thermonectus marmoratus. In both cases, we find that multiple aspects of ocular formation are disrupted, including lens facet organization and optics as well as photoreceptor morphogenesis. Together, our findings support the possibility of a generalized role for SCs in arthropod ommatidial form and function and introduces Cut as a central player in mediating this role.
Keywords: compound eyes; cone cells; conserved gene networks; invertebrates; optics; semper cells; visual system development.
Copyright © 2023 Rathore, Meece, Charlton-Perkins, Cook and Buschbeck.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


Similar articles
-
Exploring the molecular makeup of support cells in insect camera eyes.BMC Genomics. 2023 Nov 22;24(1):702. doi: 10.1186/s12864-023-09804-5. BMC Genomics. 2023. PMID: 37993800 Free PMC article.
-
Semper's cells in the insect compound eye: Insights into ocular form and function.Dev Biol. 2021 Nov;479:126-138. doi: 10.1016/j.ydbio.2021.07.015. Epub 2021 Jul 31. Dev Biol. 2021. PMID: 34343526 Free PMC article. Review.
-
Exploring the molecular makeup of support cells in insect camera eyes.bioRxiv [Preprint]. 2023 Jul 20:2023.07.19.549729. doi: 10.1101/2023.07.19.549729. bioRxiv. 2023. Update in: BMC Genomics. 2023 Nov 22;24(1):702. doi: 10.1186/s12864-023-09804-5. PMID: 37503285 Free PMC article. Updated. Preprint.
-
Embryonic development of the larval eyes of the Sunburst Diving Beetle, Thermonectus marmoratus (Insecta: Dytiscidae): a morphological study.Evol Dev. 2016 Jul;18(4):216-28. doi: 10.1111/ede.12192. Evol Dev. 2016. PMID: 27402568
-
Lens crystallins of invertebrates--diversity and recruitment from detoxification enzymes and novel proteins.Eur J Biochem. 1996 Feb 1;235(3):449-65. doi: 10.1111/j.1432-1033.1996.00449.x. Eur J Biochem. 1996. PMID: 8654388 Review.
Cited by
-
Cataract induction in an arthropod reveals how lens crystallins contribute to the formation of biological glass.PLoS One. 2025 Jun 11;20(6):e0325229. doi: 10.1371/journal.pone.0325229. eCollection 2025. PLoS One. 2025. PMID: 40498792 Free PMC article.
-
Osmosis as nature's method for establishing optical alignment.Curr Biol. 2024 Apr 8;34(7):1569-1575.e3. doi: 10.1016/j.cub.2024.02.052. Epub 2024 Mar 20. Curr Biol. 2024. PMID: 38513653 Free PMC article.
-
Exploring the molecular makeup of support cells in insect camera eyes.BMC Genomics. 2023 Nov 22;24(1):702. doi: 10.1186/s12864-023-09804-5. BMC Genomics. 2023. PMID: 37993800 Free PMC article.
References
-
- Belušič G. (2011). “ERG in Drosophila,” in Electroretinograms. Editor Belušič G. (London: IntecOpen; ). 10.5772/21747 - DOI
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials