

This is a preprint.
Interferon ε restricts Zika virus infection in the female reproductive tract
- PMID: 37066223
- PMCID: PMC10104157
- DOI: 10.1101/2023.04.06.535968
Interferon ε restricts Zika virus infection in the female reproductive tract
Update in
-
Interferon ɛ restricts Zika virus infection in the female reproductive tract.PNAS Nexus. 2023 Oct 25;2(11):pgad350. doi: 10.1093/pnasnexus/pgad350. eCollection 2023 Nov. PNAS Nexus. 2023. PMID: 37954158 Free PMC article.
Abstract
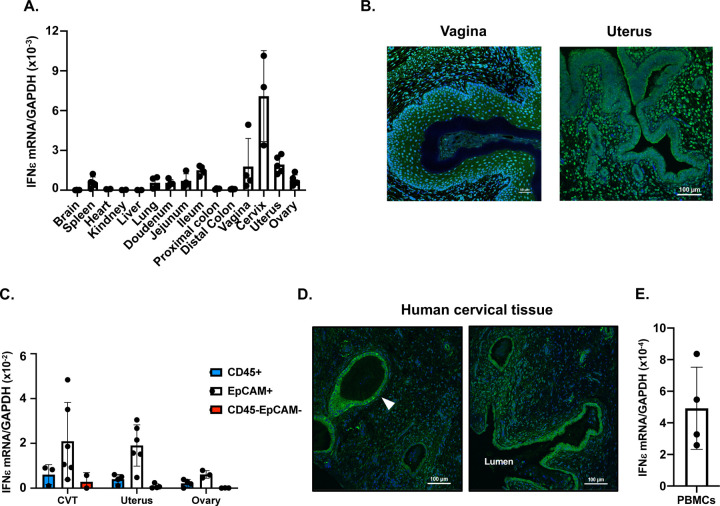
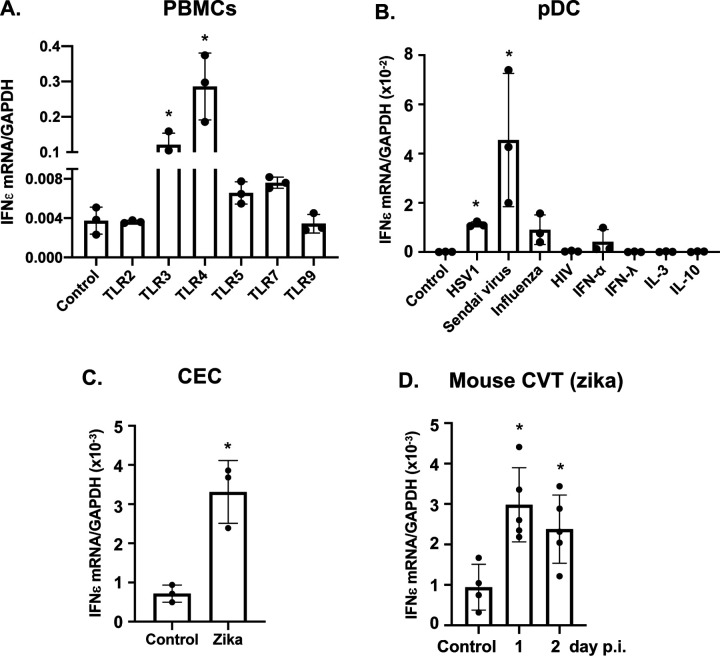
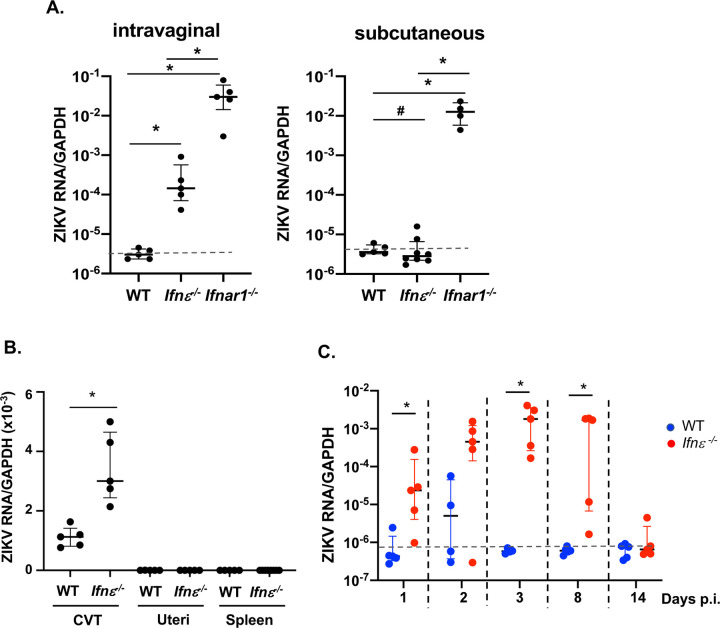
Interferon ε (IFNε) is a unique type I IFN that has been implicated in host defense against sexually transmitted infections (STIs). Zika virus (ZIKV), an emerging pathogen, can infect the female reproductive tract (FRT) and cause devastating diseases, particularly in pregnant women. How IFNε contributes to protection against ZIKV infection in vivo is unknown. Here, we show that IFNε plays a critical role in host protection against vaginal ZIKV infection in mice. We found that IFNε was expressed not only by epithelial cells in the FRT, but also by certain immune and other cells at baseline or after exposure to viruses or specific TLR agonists. IFNε-deficient mice exhibited abnormalities in the epithelial border and underlying tissue in the cervicovaginal tract, and these defects were associated with increased susceptibility to vaginal, but not subcutaneous ZIKV infection. IFNε-deficiency resulted in an increase in magnitude, duration, and depth of ZIKV infection in the FRT. Critically, intravaginal administration of recombinant IFNε protected Ifnε-/- mice and highly susceptible Ifnar1-/- mice against vaginal ZIKV infection, indicating that IFNε was sufficient to provide protection even in the absence of signals from other type I IFNs and in an IFNAR1-independent manner. Our findings reveal a potentially critical role for IFNε in mediating protection against transmission of ZIKV in the context of sexual contact.
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures



Similar articles
-
Interferon ɛ restricts Zika virus infection in the female reproductive tract.PNAS Nexus. 2023 Oct 25;2(11):pgad350. doi: 10.1093/pnasnexus/pgad350. eCollection 2023 Nov. PNAS Nexus. 2023. PMID: 37954158 Free PMC article.
-
Constitutive expression and distinct properties of IFN-epsilon protect the female reproductive tract from Zika virus infection.PLoS Pathog. 2023 Mar 10;19(3):e1010843. doi: 10.1371/journal.ppat.1010843. eCollection 2023 Mar. PLoS Pathog. 2023. PMID: 36897927 Free PMC article.
-
Zika Virus Replicates in the Vagina of Mice with Intact Interferon Signaling.J Virol. 2022 Sep 28;96(18):e0121922. doi: 10.1128/jvi.01219-22. Epub 2022 Aug 30. J Virol. 2022. PMID: 36040178 Free PMC article.
-
Running interference: Interplay between Zika virus and the host interferon response.Cytokine. 2019 Jul;119:7-15. doi: 10.1016/j.cyto.2019.02.009. Epub 2019 Mar 8. Cytokine. 2019. PMID: 30856603 Review.
-
Zika Virus in the Male Reproductive Tract.Viruses. 2018 Apr 16;10(4):198. doi: 10.3390/v10040198. Viruses. 2018. PMID: 29659541 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases