

This is a preprint.
The Chromatin Landscape of Pathogenic Transcriptional Cell States in Rheumatoid Arthritis
- PMID: 37066336
- PMCID: PMC10104143
- DOI: 10.1101/2023.04.07.536026
The Chromatin Landscape of Pathogenic Transcriptional Cell States in Rheumatoid Arthritis
Update in
-
The chromatin landscape of pathogenic transcriptional cell states in rheumatoid arthritis.Nat Commun. 2024 May 31;15(1):4650. doi: 10.1038/s41467-024-48620-7. Nat Commun. 2024. PMID: 38821936 Free PMC article.
Abstract
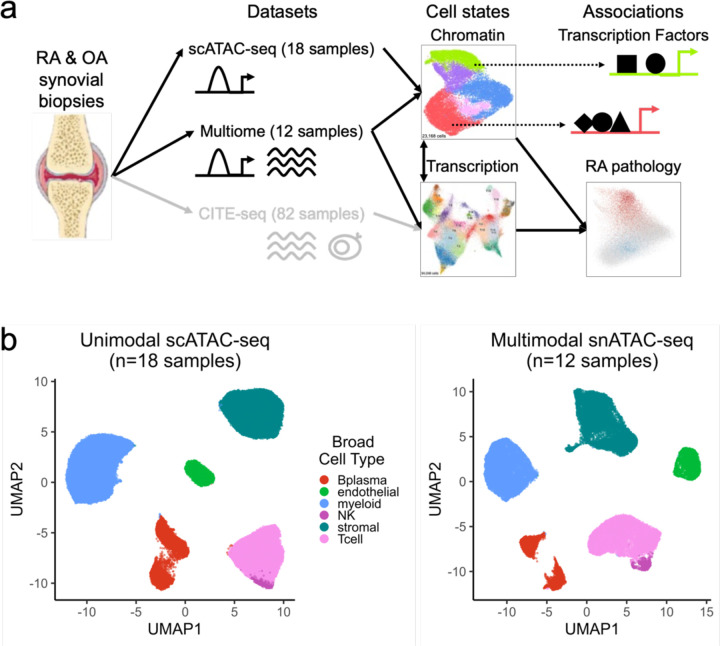
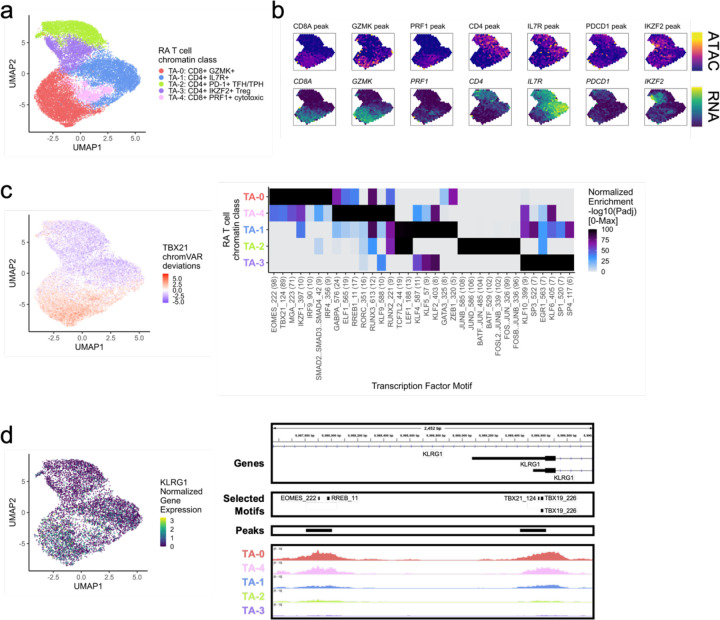
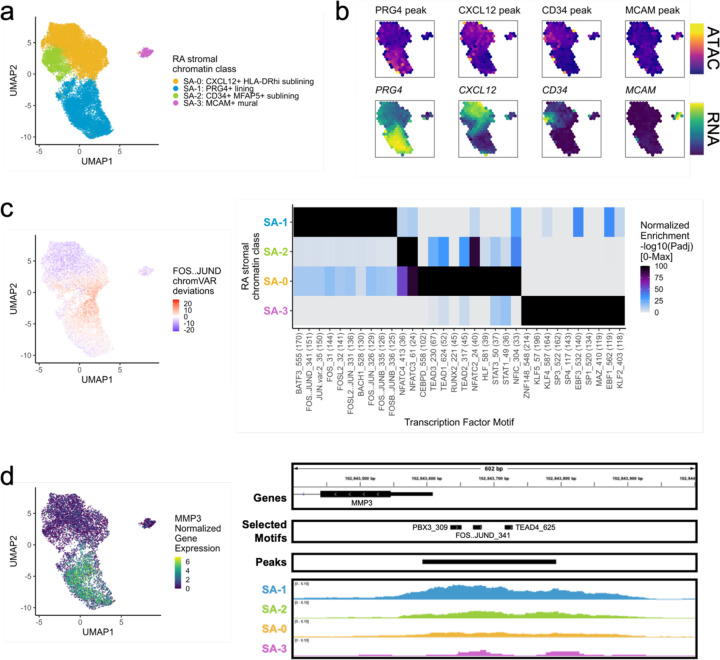
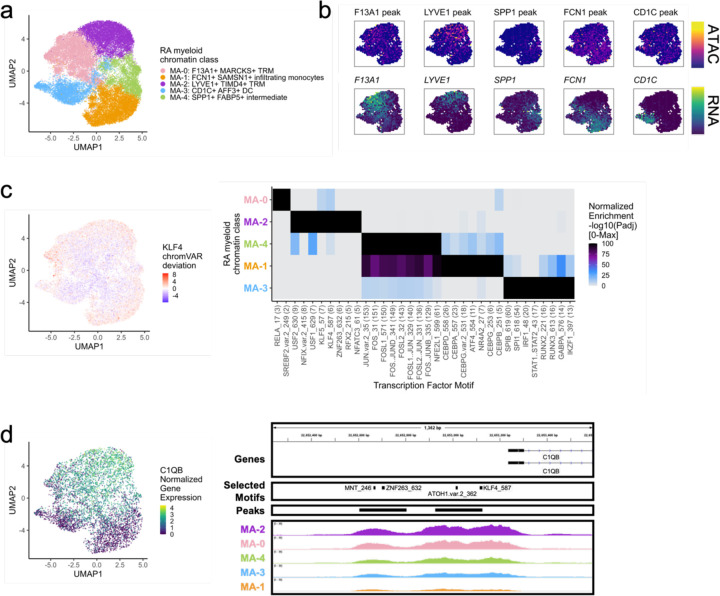
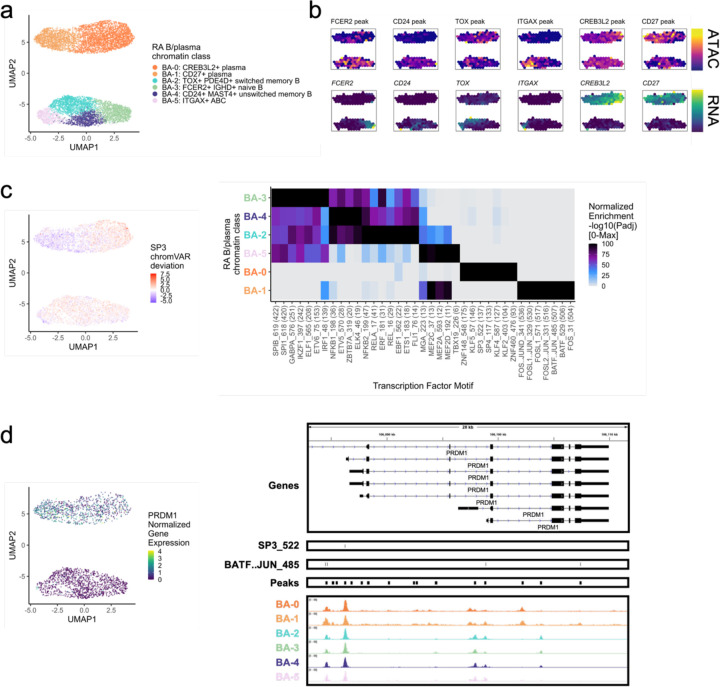
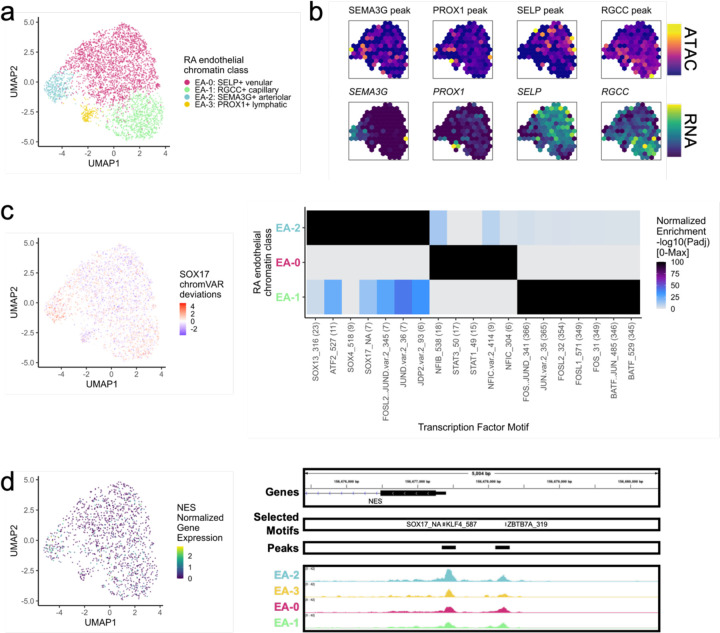
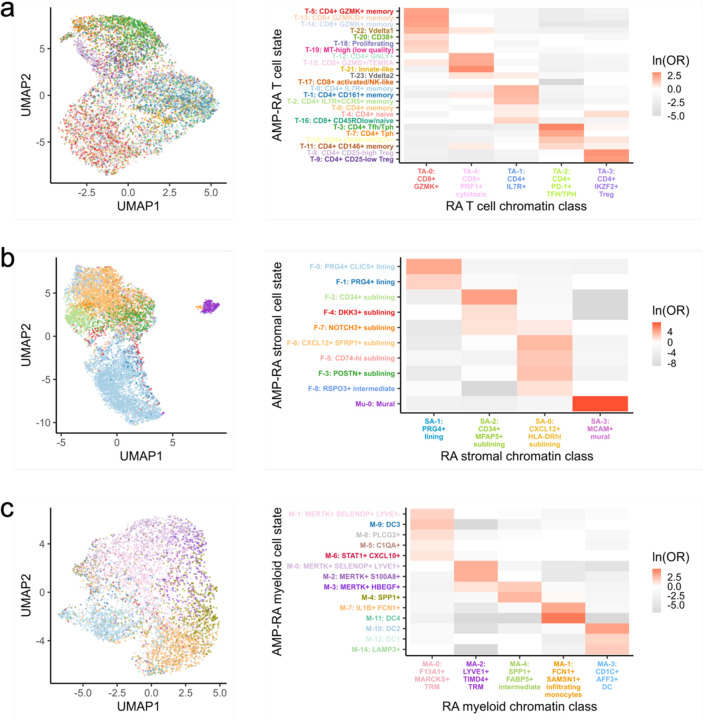
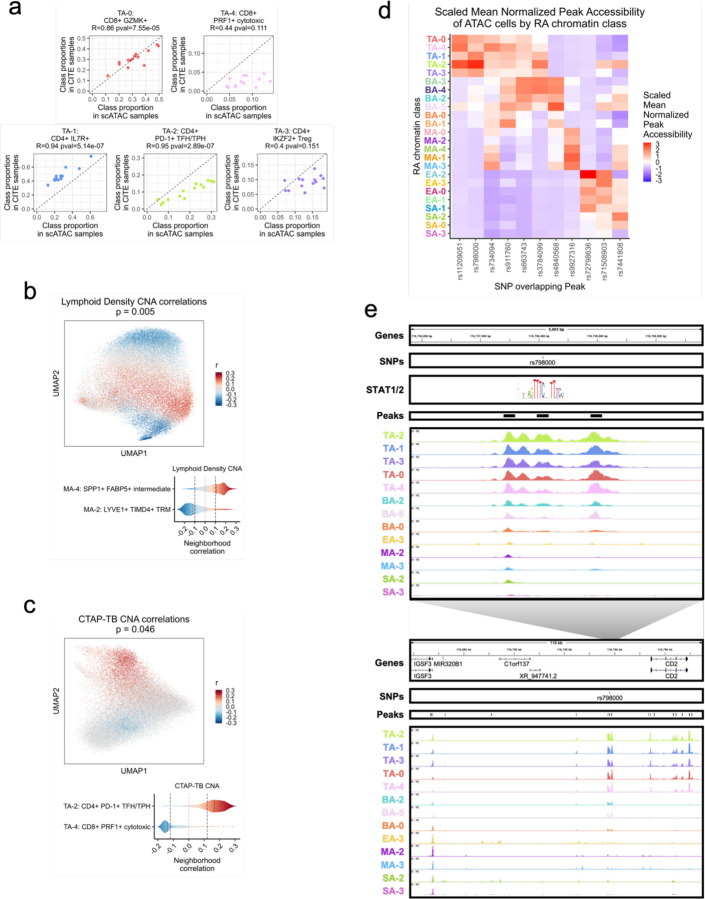
Synovial tissue inflammation is the hallmark of rheumatoid arthritis (RA). Recent work has identified prominent pathogenic cell states in inflamed RA synovial tissue, such as T peripheral helper cells; however, the epigenetic regulation of these states has yet to be defined. We measured genome-wide open chromatin at single cell resolution from 30 synovial tissue samples, including 12 samples with transcriptional data in multimodal experiments. We identified 24 chromatin classes and predicted their associated transcription factors, including a CD8+ GZMK+ class associated with EOMES and a lining fibroblast class associated with AP-1. By integrating an RA tissue transcriptional atlas, we found that the chromatin classes represented 'superstates' corresponding to multiple transcriptional cell states. Finally, we demonstrated the utility of this RA tissue chromatin atlas through the associations between disease phenotypes and chromatin class abundance as well as the nomination of classes mediating the effects of putatively causal RA genetic variants.
Conflict of interest statement
Competing Interests S.R. is a founder for Mestag Therapeutics, a scientific advisor for Janssen and Pfizer, and a consultant for Gilead. D.A.R. reports personal fees from Pfizer, Janssen, Merck, GlaxoSmithKline, AstraZeneca, Scipher Medicine, HiFiBio, and Bristol-Myers Squibb, and grant support from Merck, Janssen, and Bristol-Myers Squibb outside the submitted work. D.A.R. is a co-inventor on the patent for Tph cells as a biomarker of autoimmunity.
Figures
References
Publication types
Grants and funding
- UH2 AR067685/AR/NIAMS NIH HHS/United States
- UM2 AR067678/AR/NIAMS NIH HHS/United States
- UC2 AR081023/AR/NIAMS NIH HHS/United States
- T32 HG002295/HG/NHGRI NIH HHS/United States
- T32 AR007530/AR/NIAMS NIH HHS/United States
- UH2 AR067681/AR/NIAMS NIH HHS/United States
- UH2 AR067688/AR/NIAMS NIH HHS/United States
- UH2 AR067689/AR/NIAMS NIH HHS/United States
- UH2 AR067694/AR/NIAMS NIH HHS/United States
- K08 AR077037/AR/NIAMS NIH HHS/United States
- UH2 AR067679/AR/NIAMS NIH HHS/United States
- R01 AR078769/AR/NIAMS NIH HHS/United States
- UH2 AR067690/AR/NIAMS NIH HHS/United States
- U01 HG009379/HG/NHGRI NIH HHS/United States
- UH2 AR067677/AR/NIAMS NIH HHS/United States
- UH2 AR067676/AR/NIAMS NIH HHS/United States
- UH2 AR067691/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials