

RAF1 deficiency causes a lethal syndrome that underscores RTK signaling during embryogenesis
- PMID: 37066513
- PMCID: PMC10165362
- DOI: 10.15252/emmm.202217078
RAF1 deficiency causes a lethal syndrome that underscores RTK signaling during embryogenesis
Abstract
Somatic and germline gain-of-function point mutations in RAF, one of the first oncogenes to be discovered in humans, delineate a group of tumor-prone syndromes known as the RASopathies. In this study, we document the first human phenotype resulting from the germline loss-of-function of the proto-oncogene RAF1 (a.k.a. CRAF). In a consanguineous family, we uncovered a homozygous p.Thr543Met variant segregating with a neonatal lethal syndrome with cutaneous, craniofacial, cardiac, and limb anomalies. Structure-based prediction and functional tests using human knock-in cells showed that threonine 543 is essential to: (i) ensure RAF1's stability and phosphorylation, (ii) maintain its kinase activity toward substrates of the MAPK pathway, and (iii) protect from stress-induced apoptosis mediated by ASK1. In Xenopus embryos, mutant RAF1T543M failed to phenocopy the effects of normal and overactive FGF/MAPK signaling, confirming its hypomorphic activity. Collectively, our data disclose the genetic and molecular etiology of a novel lethal syndrome with progeroid features, highlighting the importance of RTK signaling for human development and homeostasis.
Keywords: Xenopus; ASK1; ERK; RAF1; RASopathy.
© 2023 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

- A
Pedigree of the Turkish consanguineous family reporting the neonatal progeroid syndrome. Square, male; circle, female; black shading, affected individuals; small triangle, aborted; double lines, consanguineous marriages; diagonal line, deceased.
- B
Pictures of affected proband (III:2) at 50 days showing general progeroid features. Note subcutaneous lipoatrophy, severe hypotonia, distinctive craniofacial features with absence of external ears (insets), cleft hands and feet with oligodactyly.
- C
Schematic of the RAF1 protein structure showing the three conserved regions (CRs). p.T543M is located in the catalytic kinase domain (CR3).
- D
Multiple sequence alignment showing conservation of T543 across species and among RAF family members.
- E
RAF1WT or RAF1T543M were transiently overexpressed in HEK293T cells for 48 h, followed by 16 h of serum starvation and EGF stimulation for 15 min. Phos‐tag™ gel showing global levels of RAF1 phosphorylation and corresponding Western blot showing phosphorylation levels of Ser259 and Ser338 residues and total RAF1.
- F, G
All the RAF1 constructs were transiently overexpressed in HEK293T cells as described in (E). Western blotting showing (F) phosphorylated MEK1/2, total MEK1/2 and total RAF1 and (G) phosphorylated ERK1/2, total ERK1/2 and total RAF1.

- A
Genetic strategy. Combination of exome sequencing and homozygosity mapping. resulted in four candidate genes. Germline homozygous variants in PLIN2, ABL1, and C16orf93 were filtered out due to their low phylogenetic conservation and low predicted deleterious potential.
- B
Sanger sequencing chromatogram showing variant c.1628C>T (p.T543M) segregating with disease: homozygous in the proband (III:2), heterozygous in the parents (II:2 and II:3) and one sister (III:4) and absent in two other siblings (III:3 and III:6).
- C
Schematic representation of ERK1/2 MAP Kinase pathway.
- D
Quantification of phosphorylated ERK1/2, normalized to WT RAF1 in three independent Western blots (Error bars indicate mean ± SEM. Ordinary one‐way ANOVA).

- A
Western blotting showing phosphorylated Erk1/2, total Erk1/2 and total Raf1 from Xenopus laevis embryos harvested at stage 28.
- B
Quantification of phosphorylated Erk1/2, normalized to WT Raf1 in three independent Western blots (Error bars indicate mean ± SEM. Ordinary one‐way ANOVA).
- C
Representative images illustrating the effect of WT or mutants RAF1 mRNA injection into Xenopus embryos, at stage 28.
- D
WISH performed on stage 10.5 Xenopus embryos injected with WT or mutants RAF1 mRNA. Xbra (blue staining) marking mesoderm induction. Vegetal (top), animal (middle) and lateral (bottom) views.
- E
WISH performed on stage 28 Xenopus embryos injected with WT or mutants RAF1 mRNA. Blue staining reveals Xbra or N‐tubulin markers depicting ectopic mesoderm induction, neural differentiation and brain posterization.
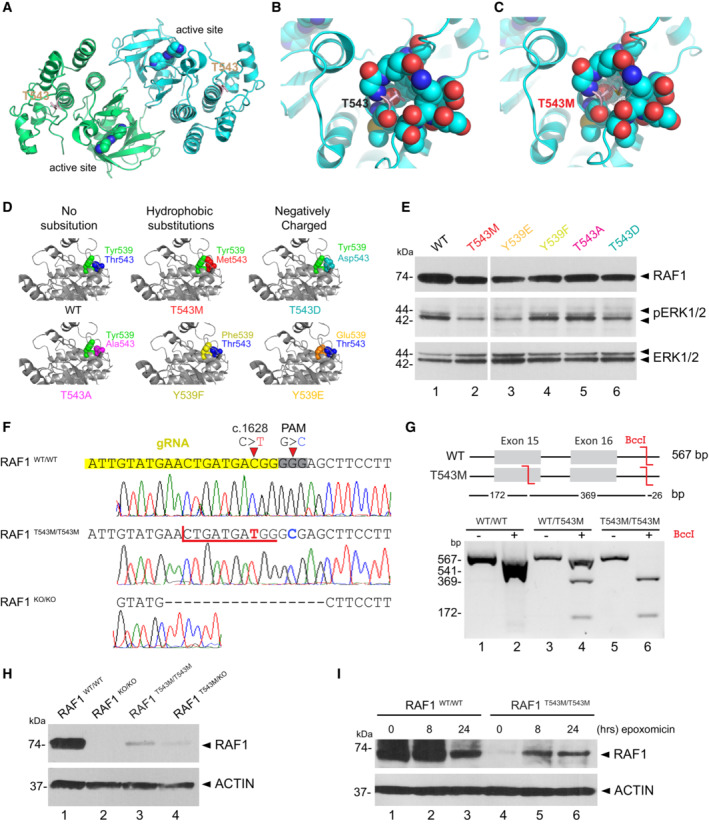
- A
RAF1 homodimer (PDB:3OMV) showing the location of Thr543 (drawn as ball‐and‐stick) on helical subdomain. Active site is occupied by an inhibitor drawn as spheres.
- B
Positioning of Thr543 in the pocket.
- C
Met543 in the pocket. The Met543 side chain clashes despite being modeled in the least sterically hindered rotamer.
- D
Predicted side chain interaction of Thr543 and Tyr539 variants.
- E
Western blot downstream signaling potential of structural mutants that induce steric (p.T543M) or electrostatic (p.Y539E or p.T543D) interference.
- F
RAF1T543M and RAF1KO/KO HEK293T cells were generated by CRISPR‐Cas9. Top—bottom: Sanger‐sequencing of RAF1 in WT HEK293T cells; a RAF1T543M clone showing the desired c.1628C>T (p.T543M) mutation and a synonymous PAM‐disrupting c.1632G>C; a RAF1KO/KO clone with a homozygous 17 bp deletion.
- G
Confirmation of a clonal RAF1T543M line by restriction‐digest of RAF1 exons 15–16 from gDNA of the genotypes shown with BccI. c.1628C>T introduces a BccI restriction site. The RAF1WT/T543M heterozygote genotype was obtained from the patient's father.
- H
Western Blot for endogenous RAF1 in HEK293T cell lines of genotypes indicated, at steady state.
- I
Western‐blot showing that 8 and 24 h of epoxomicin treatment is sufficient to partially rescue RAF1 in RAF1T543M/T543M cells. ACTIN serves as loading control.

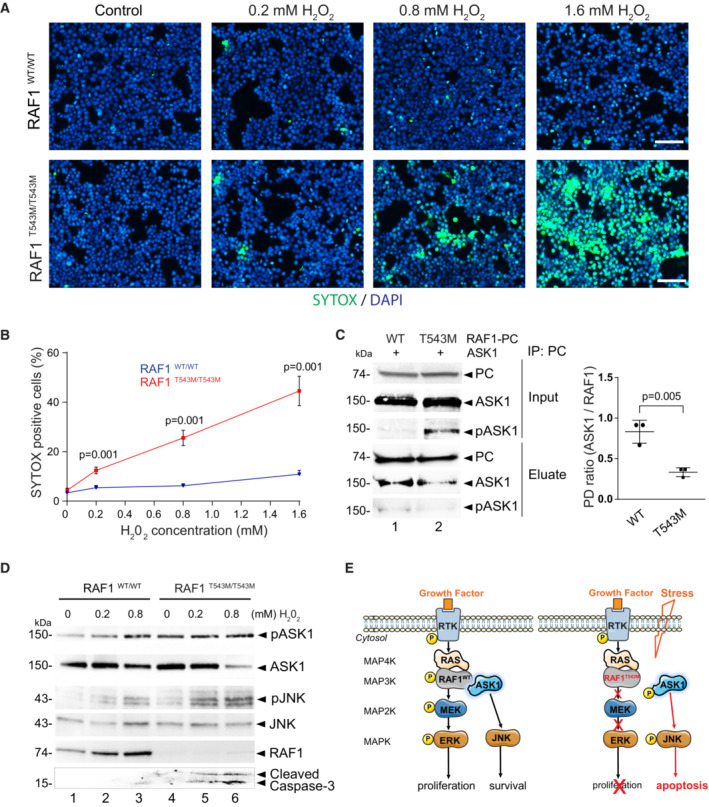
- A
Representative immunofluorescence images of 0.2, 0.8, and 1.6 mM H2O2 treated RAF1T543M/T543M and RAF1WT/WT HEK293T cells. Green: SYTOX dead cell stain. Blue: DAPI nuclei stain. Scale bar, 100 μm.
- B
Dose‐dependent SYTOX signal of RAF1T543M/T543M and RAF1WT/WT HEK293T cells incubated with increasing H2O2 concentrations for 2 h (n = 8 from two independent experiments, error bars indicate mean ± SEM. Two‐tailed Student's t‐test).
- C
PC tagged RAF1WT or RAF1T543M were transiently overexpressed along with ASK1 in HEK293T cells for 48 h, immunoprecipitated and blotted for ASK1. Ratio of ASK1 to RAF1 from the pulldown (PD) was quantified in three independent Western blots (Error bars indicate mean ± SEM. Ordinary one‐way ANOVA).
- D
Western blot documenting endogenous pASK1, pJNK levels and cleaved Caspase 3 following H2O2‐induced stress in isogenic HEK293T cells expressing endogenous RAF1T543M/T543M or RAF1WT. Note that pASK1 is increased without H2O2 treatment in RAF1T543M/T543M compared with RAF1WT.
- E
Proposed model of the negative impact of RAF1T543M/T543M on proliferation owing to impaired MAPK/ERK transduction and increased apoptosis due to de‐repression of ASK1 signaling.

References
-
- Aoki Y, Niihori T, Inoue S‐I, Matsubara Y (2016) Recent advances in RASopathies. J Hum Genet 61: 33–39 - PubMed
-
- Baccarini M (2005) Second nature: biological functions of the Raf‐1 ‘kinase’. FEBS Lett 579: 3271–3277 - PubMed
-
- Bentires‐Alj M, Kontaridis MI, Neel BG (2006) Stops along the RAS pathway in human genetic disease. Nat Med 12: 283–285 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
