

Dimeric p53 Mutant Elicits Unique Tumor-Suppressive Activities through an Altered Metabolic Program
- PMID: 37067911
- PMCID: PMC10164062
- DOI: 10.1158/2159-8290.CD-22-0872
Dimeric p53 Mutant Elicits Unique Tumor-Suppressive Activities through an Altered Metabolic Program
Abstract
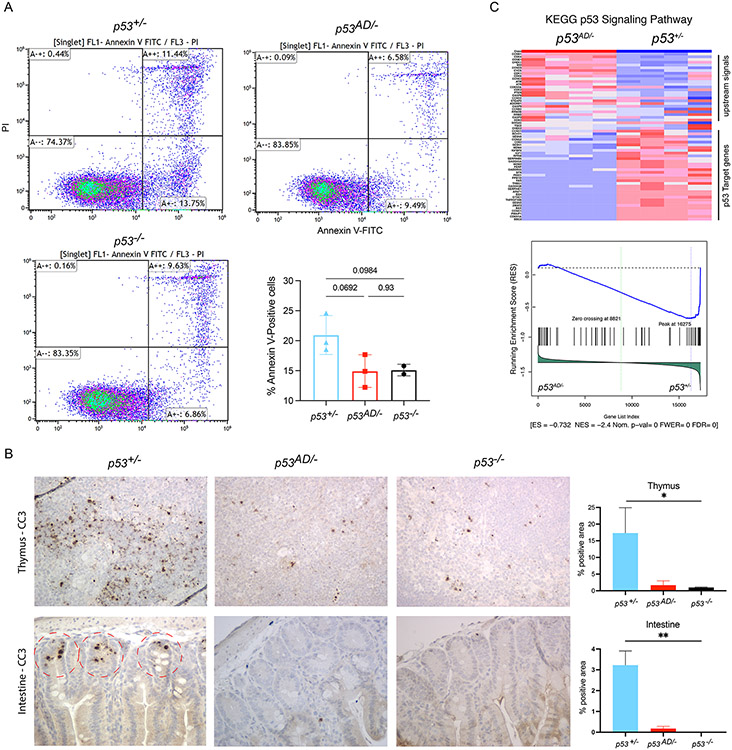
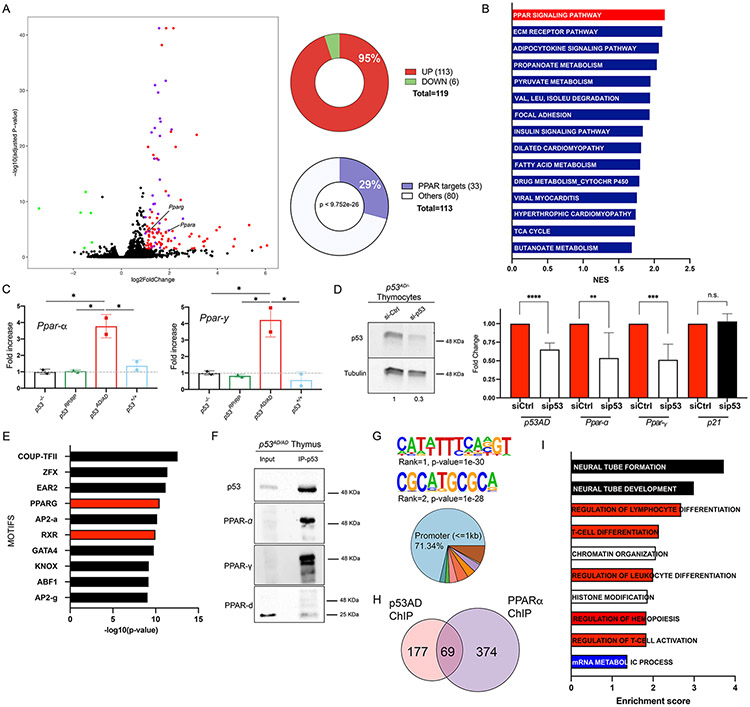
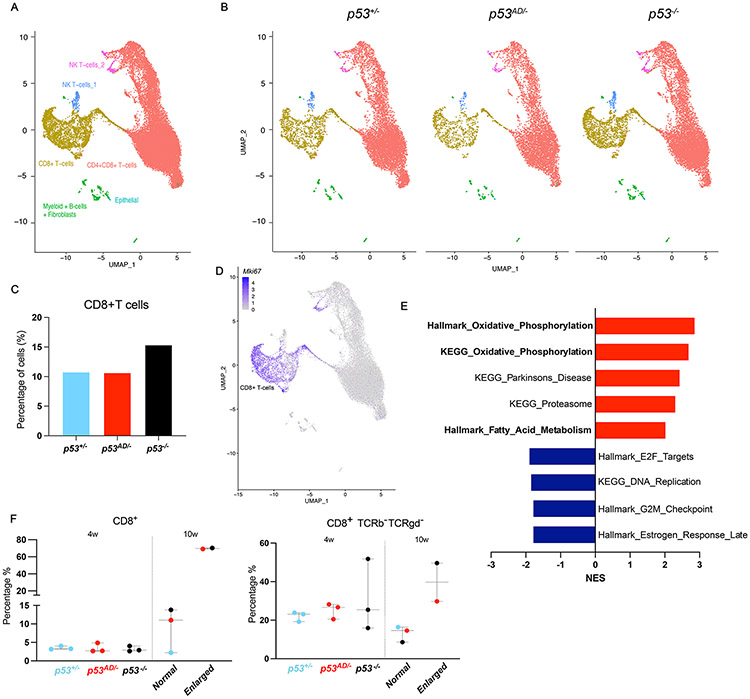
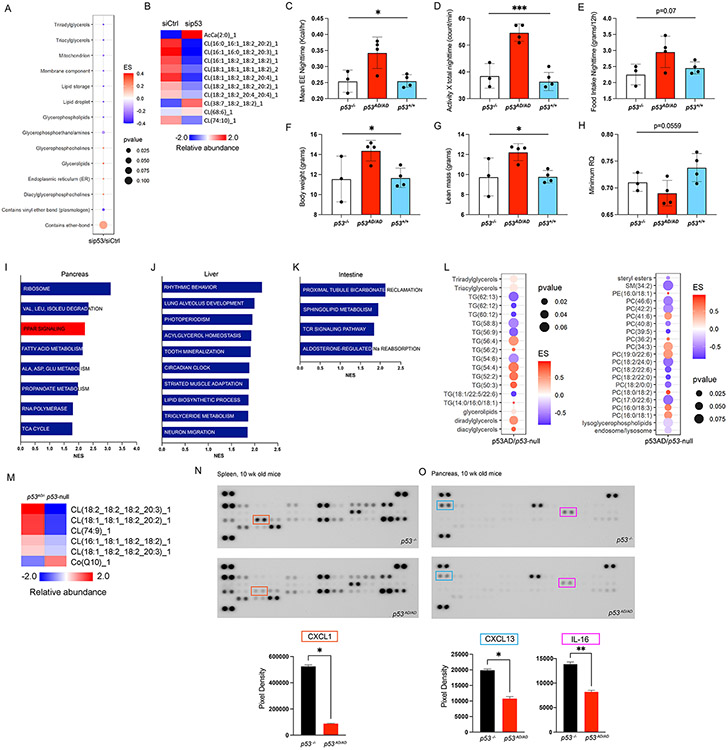
Cancer-related alterations of the p53 tetramerization domain (TD) abrogate wild-type (WT) p53 function. They result in a protein that preferentially forms monomers or dimers, which are also normal p53 states under basal cellular conditions. However, their physiologic relevance is not well understood. We have established in vivo models for monomeric and dimeric p53, which model Li-Fraumeni syndrome patients with germline p53 TD alterations. p53 monomers are inactive forms of the protein. Unexpectedly, p53 dimers conferred some tumor suppression that is not mediated by canonical WT p53 activities. p53 dimers upregulate the PPAR pathway. These activities are associated with lower prevalence of thymic lymphomas and increased CD8+ T-cell differentiation. Lymphomas derived from dimeric p53 mice show cooperating alterations in the PPAR pathway, further implicating a role for these activities in tumor suppression. Our data reveal novel functions for p53 dimers and support the exploration of PPAR agonists as therapies.
Significance: New mouse models with TP53R342P (monomer) or TP53A347D (dimer) mutations mimic Li-Fraumeni syndrome. Although p53 monomers lack function, p53 dimers conferred noncanonical tumor-suppressive activities. We describe novel activities for p53 dimers facilitated by PPARs and propose these are "basal" p53 activities. See related commentary by Stieg et al., p. 1046. See related article by Choe et al., p. 1250. This article is highlighted in the In This Issue feature, p. 1027.
©2023 American Association for Cancer Research.
Conflict of interest statement
Conflict-of-interest statement:
The authors declare no potential conflicts of interest.
Figures

Comment in
-
Li-Fraumeni Syndrome-Associated Dimer-Forming Mutant p53 Promotes Transactivation-Independent Mitochondrial Cell Death.Cancer Discov. 2023 May 4;13(5):1250-1273. doi: 10.1158/2159-8290.CD-22-0882. Cancer Discov. 2023. PMID: 37067901 Free PMC article.
-
p53 Oligomerization Domain Mutants: A New Class of Mutants That Retain "License to Kill".Cancer Discov. 2023 May 4;13(5):1046-1048. doi: 10.1158/2159-8290.CD-23-0200. Cancer Discov. 2023. PMID: 37139723
Comment on
-
Li-Fraumeni Syndrome-Associated Dimer-Forming Mutant p53 Promotes Transactivation-Independent Mitochondrial Cell Death.Cancer Discov. 2023 May 4;13(5):1250-1273. doi: 10.1158/2159-8290.CD-22-0882. Cancer Discov. 2023. PMID: 37067901 Free PMC article.
References
-
- Riley T, Sontag E, Chen P, and Levine A, Transcriptional control of human p53-regulated genes. Nat Rev Mol Cell Biol, 2008. 9(5): p. 402–12. - PubMed
-
- el-Deiry WS, Kern SE, Pietenpol JA, Kinzler KW, and Vogelstein B, Definition of a consensus binding site for p53. Nat Genet, 1992. 1(1): p. 45–9. - PubMed
-
- Weinberg RL, Veprintsev DB, and Fersht AR, Cooperative binding of tetrameric p53 to DNA. J Mol Biol, 2004. 341(5): p. 1145–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
