

Ether phospholipids are required for mitochondrial reactive oxygen species homeostasis
- PMID: 37069167
- PMCID: PMC10110566
- DOI: 10.1038/s41467-023-37924-9
Ether phospholipids are required for mitochondrial reactive oxygen species homeostasis
Abstract
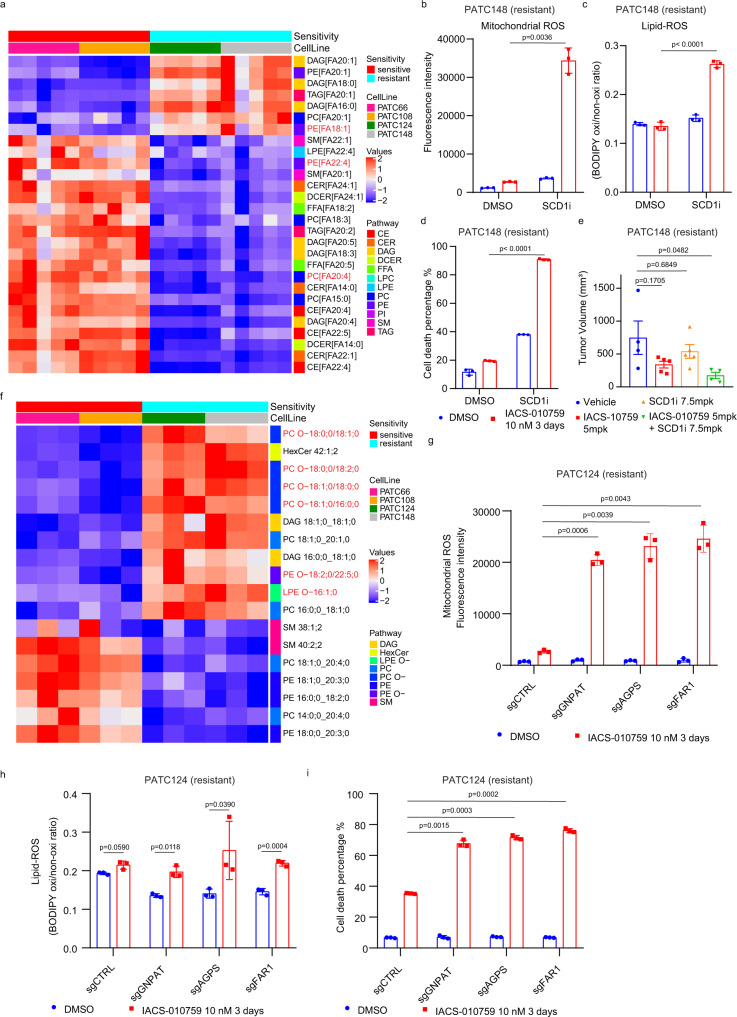
Mitochondria are hubs where bioenergetics, redox homeostasis, and anabolic metabolism pathways integrate through a tightly coordinated flux of metabolites. The contributions of mitochondrial metabolism to tumor growth and therapy resistance are evident, but drugs targeting mitochondrial metabolism have repeatedly failed in the clinic. Our study in pancreatic ductal adenocarcinoma (PDAC) finds that cellular and mitochondrial lipid composition influence cancer cell sensitivity to pharmacological inhibition of electron transport chain complex I. Profiling of patient-derived PDAC models revealed that monounsaturated fatty acids (MUFAs) and MUFA-linked ether phospholipids play a critical role in maintaining ROS homeostasis. We show that ether phospholipids support mitochondrial supercomplex assembly and ROS production; accordingly, blocking de novo ether phospholipid biosynthesis sensitized PDAC cells to complex I inhibition by inducing mitochondrial ROS and lipid peroxidation. These data identify ether phospholipids as a regulator of mitochondrial redox control that contributes to the sensitivity of PDAC cells to complex I inhibition.
© 2023. The Author(s).
Conflict of interest statement
C.A.L. has received consulting fees from Astellas Pharmaceuticals and Odyssey Therapeutics, and is an inventor on patents pertaining to Kras regulated metabolic pathways, redox control pathways in cancer, and targeting the GOT1-pathway as a therapeutic approach. The remaining authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
- P50 CA221707/CA/NCI NIH HHS/United States
- P01CA117969/NH/NIH HHS/United States
- R01 CA248160/CA/NCI NIH HHS/United States
- R01 CA244931/CA/NCI NIH HHS/United States
- R01 CA258917/CA/NCI NIH HHS/United States
- S10 OD012304/OD/NIH HHS/United States
- R01 CA214793/CA/NCI NIH HHS/United States
- P30 CA016672/CA/NCI NIH HHS/United States
- P01CA117969/CA/NCI NIH HHS/United States
- P50CA221707/CA/NCI NIH HHS/United States
- P01 CA117969/CA/NCI NIH HHS/United States
- S10OD012304-01/NH/NIH HHS/United States
- R01CA248160/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Research Materials
