

An asymmetric structure of bacterial TrpRS supports the half-of-the-sites catalytic mechanism and facilitates antimicrobial screening
- PMID: 37070195
- PMCID: PMC10201369
- DOI: 10.1093/nar/gkad278
An asymmetric structure of bacterial TrpRS supports the half-of-the-sites catalytic mechanism and facilitates antimicrobial screening
Abstract
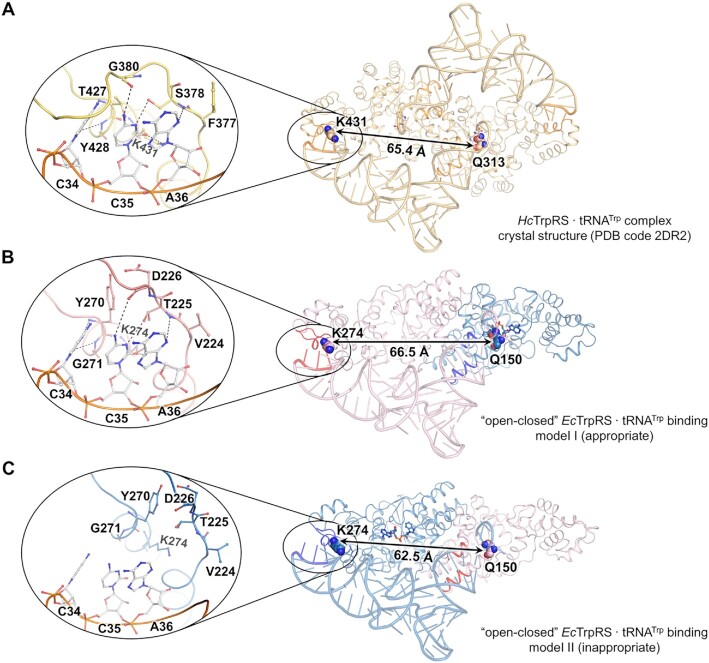
Tryptophanyl-tRNA synthetase (TrpRS) links tryptophan to tRNATrp, thereby playing an indispensable role in protein translation. Unlike most class I aminoacyl-tRNA synthetases (AARSs), TrpRS functions as a homodimer. Herein, we captured an 'open-closed' asymmetric structure of Escherichia coli TrpRS (EcTrpRS) with one active site occupied by a copurified intermediate product and the other remaining empty, providing structural evidence for the long-discussed half-of-the-sites reactivity of bacterial TrpRS. In contrast to its human counterpart, bacterial TrpRS may rely on this asymmetric conformation to functionally bind with substrate tRNA. As this asymmetric conformation is probably a dominant form of TrpRS purified from bacterial cells, we performed fragment screening against asymmetric EcTrpRS to support antibacterial discovery. Nineteen fragment hits were identified, and 8 of them were successfully cocrystallized with EcTrpRS. While a fragment named niraparib bound to the L-Trp binding site of the 'open' subunit, the other 7 fragments all bound to an unprecedented pocket at the interface between two TrpRS subunits. Binding of these fragments relies on residues specific to bacterial TrpRS, avoiding undesired interactions with human TrpRS. These findings improve our understanding of the catalytic mechanism of this important enzyme and will also facilitate the discovery of bacterial TrpRS inhibitors with therapeutic potential.
© The Author(s) 2023. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures







Similar articles
-
The mechanism of lineage-specific tRNA recognition by bacterial tryptophanyl-tRNA synthetase and its implications for inhibitor discovery.Nucleic Acids Res. 2025 May 22;53(10):gkaf466. doi: 10.1093/nar/gkaf466. Nucleic Acids Res. 2025. PMID: 40464690 Free PMC article.
-
Structures of tryptophanyl-tRNA synthetase II from Deinococcus radiodurans bound to ATP and tryptophan. Insight into subunit cooperativity and domain motions linked to catalysis.J Biol Chem. 2005 Sep 9;280(36):31965-73. doi: 10.1074/jbc.M501568200. Epub 2005 Jul 5. J Biol Chem. 2005. PMID: 15998643
-
A concerted tryptophanyl-adenylate-dependent conformational change in Bacillus subtilis tryptophanyl-tRNA synthetase revealed by the fluorescence of Trp92.J Mol Biol. 1996 Jul 19;260(3):446-66. doi: 10.1006/jmbi.1996.0413. J Mol Biol. 1996. PMID: 8757806
-
Substrate selection by aminoacyl-tRNA synthetases.Nucleic Acids Symp Ser. 1995;(33):40-2. Nucleic Acids Symp Ser. 1995. PMID: 8643392 Review.
-
Mammalian tryptophanyl-tRNA synthetases.Biochimie. 1993;75(12):1027-39. doi: 10.1016/0300-9084(93)90002-a. Biochimie. 1993. PMID: 7515282 Review.
Cited by
-
Mechanistic insights into the ATP-mediated and species-dependent inhibition of TrpRS by chuangxinmycin.RSC Chem Biol. 2025 May 9;6(7):1079-1088. doi: 10.1039/d5cb00060b. eCollection 2025 Jul 2. RSC Chem Biol. 2025. PMID: 40406164 Free PMC article.
-
The mechanism of lineage-specific tRNA recognition by bacterial tryptophanyl-tRNA synthetase and its implications for inhibitor discovery.Nucleic Acids Res. 2025 May 22;53(10):gkaf466. doi: 10.1093/nar/gkaf466. Nucleic Acids Res. 2025. PMID: 40464690 Free PMC article.
-
Purification and validation of asparaginyl-tRNA synthetase heterodimer with indistinguishable subunits.IUBMB Life. 2025 Feb;77(2):e70000. doi: 10.1002/iub.70000. IUBMB Life. 2025. PMID: 39994779
-
Substrate recognition by a peptide-aminoacyl-tRNA ligase.Proc Natl Acad Sci U S A. 2025 Mar 25;122(12):e2423858122. doi: 10.1073/pnas.2423858122. Epub 2025 Mar 19. Proc Natl Acad Sci U S A. 2025. PMID: 40106349 Free PMC article.
-
Unexpected Single-Ligand Occupancy and Negative Cooperativity in the SARS-CoV-2 Main Protease.J Chem Inf Model. 2024 Feb 12;64(3):892-904. doi: 10.1021/acs.jcim.3c01497. Epub 2023 Dec 5. J Chem Inf Model. 2024. PMID: 38051605 Free PMC article.
References
-
- Ibba M., Soll D.. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 2000; 69:617–650. - PubMed
-
- Jiang L., Jones J., Yang X.L.. Human diseases linked to cytoplasmic aminoacyl-tRNA synthetases. Enzymes. 2020; 48:277–319. - PubMed
-
- Cai Z., Chen B., Yu Y., Guo J., Luo Z., Cheng B., Xu J., Gu Q., Zhou H.. Design, synthesis, and proof-of-concept of triple-site inhibitors against aminoacyl-tRNA synthetases. J. Med. Chem. 2022; 65:5800–5820. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
