

Imprinted Anti-Hemagglutinin and Anti-Neuraminidase Antibody Responses after Childhood Infections of A(H1N1) and A(H1N1)pdm09 Influenza Viruses
- PMID: 37070986
- PMCID: PMC10294682
- DOI: 10.1128/mbio.00084-23
Imprinted Anti-Hemagglutinin and Anti-Neuraminidase Antibody Responses after Childhood Infections of A(H1N1) and A(H1N1)pdm09 Influenza Viruses
Abstract
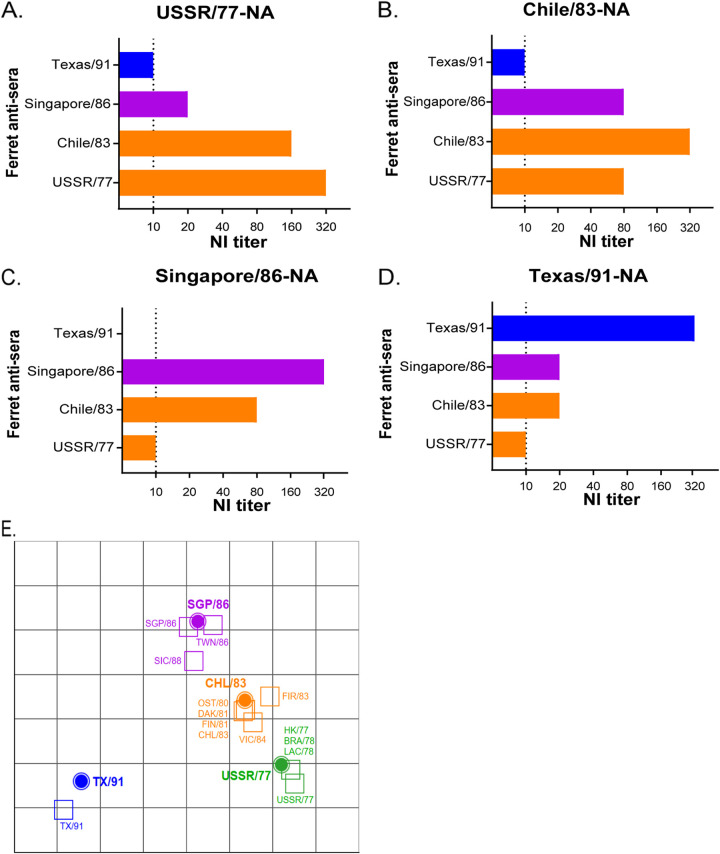
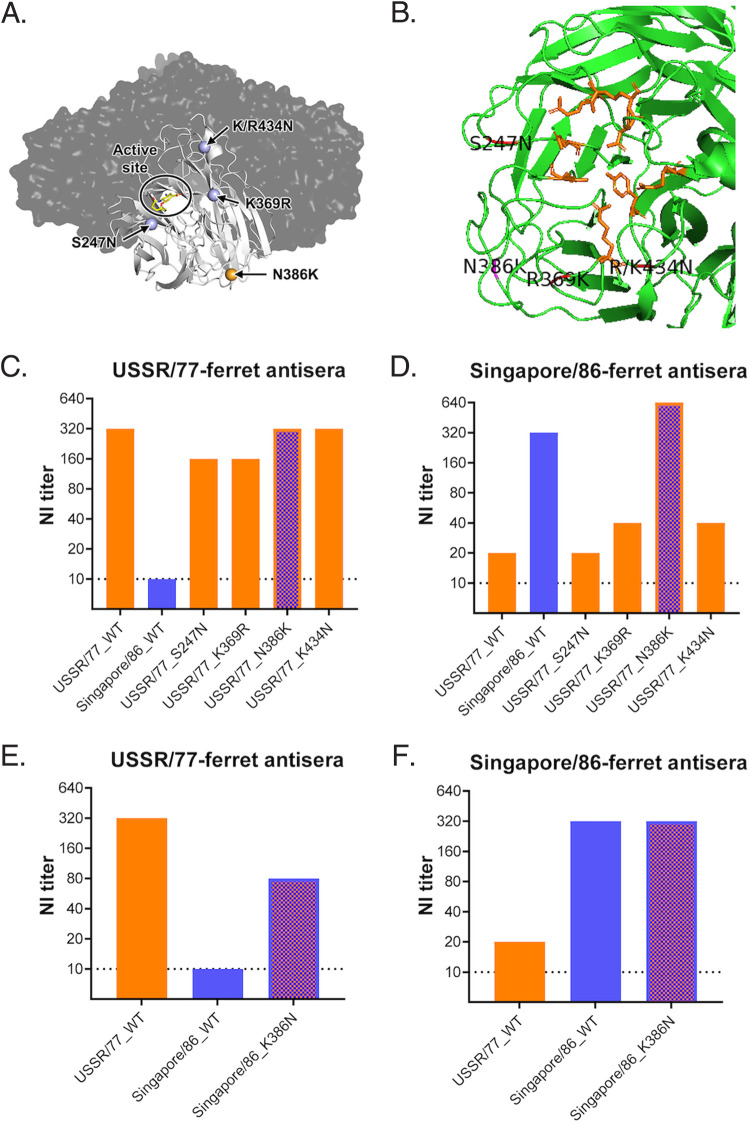
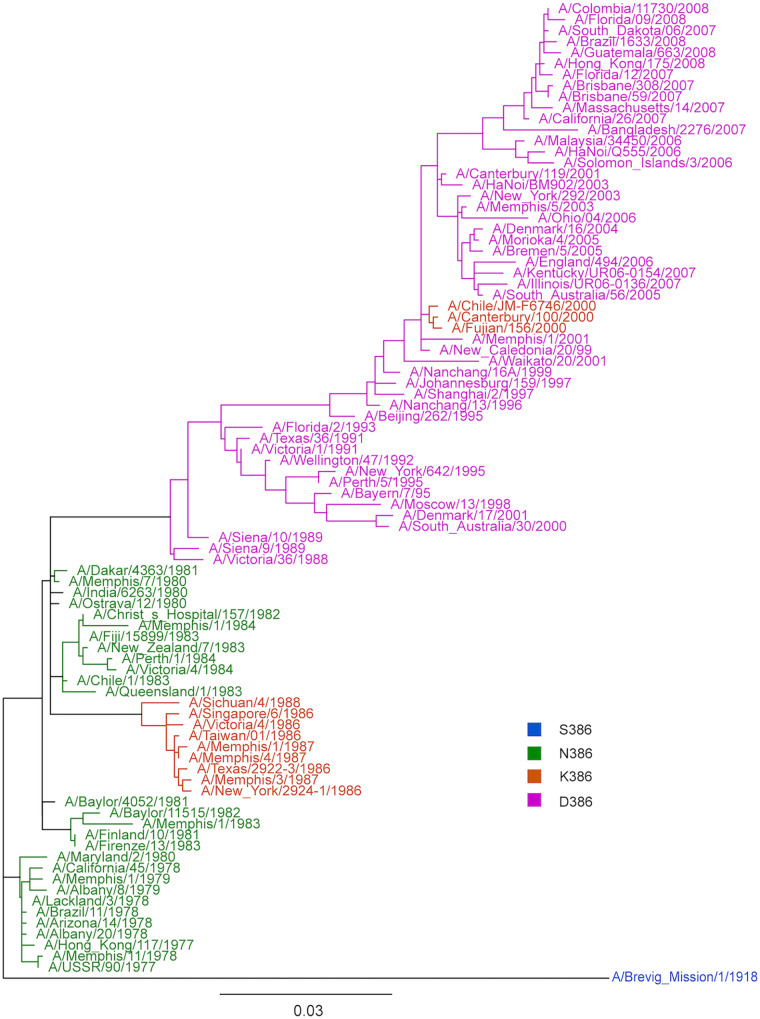
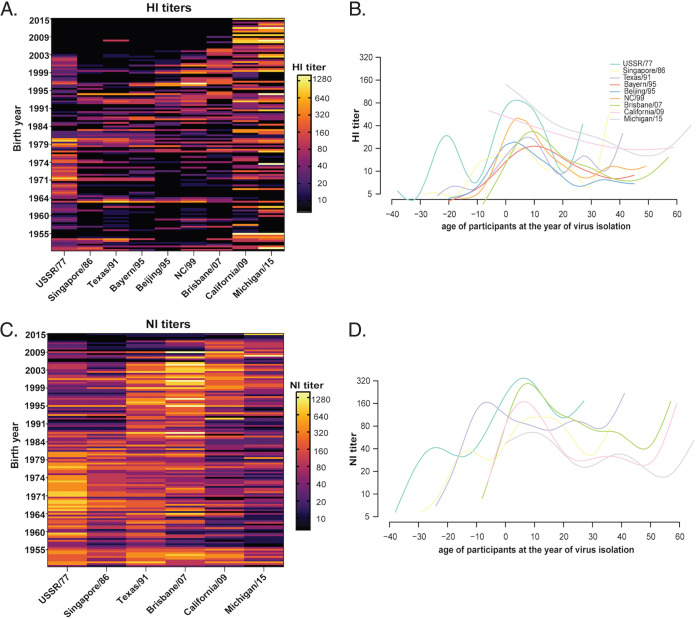
Immune imprinting is a driver known to shape the anti-hemagglutinin (HA) antibody landscape of individuals born within the same birth cohort. With the HA and neuraminidase (NA) proteins evolving at different rates under immune selection pressures, anti-HA and anti-NA antibody responses since childhood influenza virus infections have not been evaluated in parallel at the individual level. This is partly due to the limited knowledge of changes in NA antigenicity, as seasonal influenza vaccines have focused on generating neutralizing anti-HA antibodies against HA antigenic variants. Here, we systematically characterized the NA antigenic variants of seasonal A(H1N1) viruses from 1977 to 1991 and completed the antigenic profile of N1 NAs from 1977 to 2015. We identified that NA proteins of A/USSR/90/77, A/Singapore/06/86, and A/Texas/36/91 were antigenically distinct and mapped N386K as a key determinant of the NA antigenic change from A/USSR/90/77 to A/Singapore/06/86. With comprehensive panels of HA and NA antigenic variants of A(H1N1) and A(H1N1)pdm09 viruses, we determined hemagglutinin inhibition (HI) and neuraminidase inhibition (NI) antibodies from 130 subjects born between 1950 and 2015. Age-dependent imprinting was observed for both anti-HA and anti-NA antibodies, with the peak HI and NI titers predominantly detected from subjects at 4 to 12 years old during the year of initial virus isolation, except the age-independent anti-HA antibody response against A(H1N1)pdm09 viruses. More participants possessed antibodies that reacted to multiple antigenically distinct NA proteins than those with antibodies that reacted to multiple antigenically distinct HA proteins. Our results support the need to include NA proteins in seasonal influenza vaccine preparations. IMPORTANCE Seasonal influenza vaccines have aimed to generate neutralizing anti-HA antibodies for protection since licensure. More recently, anti-NA antibodies have been established as an additional correlate of protection. While HA and NA antigenic changes occurred discordantly, the anti-HA and anti-NA antibody profiles have rarely been analyzed in parallel at the individual level, due to the limited knowledge on NA antigenic changes. By characterizing NA antigenic changes of A(H1N1) viruses, we determined the anti-HA and anti-NA antibody landscape against antigenically distinct A(H1N1) and A(H1N1)pdm09 viruses using sera of 130 subjects born between 1950 and 2015. We observed age-dependent imprinting of both anti-HA and anti-NA antibodies against strains circulated during the first decade of life. A total of 67.7% (88/130) and 90% (117/130) of participants developed cross-reactive antibodies to multiple HA and NA antigens at titers ≥1:40. With slower NA antigenic changes and cross-reactive anti-NA antibody responses, including NA protein in influenza vaccine preparation may enhance vaccine efficacy.
Keywords: antigenic drift; hemagglutinin; imprinting; influenza; neuraminidase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Drift and shape-new insights into human immunity against influenza virus neuraminidase.mBio. 2023 Dec 19;14(6):e0165423. doi: 10.1128/mbio.01654-23. Epub 2023 Nov 7. mBio. 2023. PMID: 37933976 Free PMC article.
Similar articles
-
Differential antigenic imprinting effects between influenza H1N1 hemagglutinin and neuraminidase in a mouse model.J Virol. 2025 Jan 31;99(1):e0169524. doi: 10.1128/jvi.01695-24. Epub 2024 Dec 5. J Virol. 2025. PMID: 39636110 Free PMC article.
-
Targeting neuraminidase: the next frontier for broadly protective influenza vaccines.Trends Immunol. 2024 Jan;45(1):11-19. doi: 10.1016/j.it.2023.11.001. Epub 2023 Dec 15. Trends Immunol. 2024. PMID: 38103991 Free PMC article. Review.
-
Antigenic Drift of the Influenza A(H1N1)pdm09 Virus Neuraminidase Results in Reduced Effectiveness of A/California/7/2009 (H1N1pdm09)-Specific Antibodies.mBio. 2019 Apr 9;10(2):e00307-19. doi: 10.1128/mBio.00307-19. mBio. 2019. PMID: 30967460 Free PMC article.
-
Serum strain-specific or cross-reactive neuraminidase inhibiting antibodies against pandemic А/California/07/2009(H1N1) influenza in healthy volunteers.BMC Res Notes. 2015 Apr 10;8:136. doi: 10.1186/s13104-015-1086-z. BMC Res Notes. 2015. PMID: 25889924 Free PMC article.
-
Recent Advances, Approaches and Challenges in the Development of Universal Influenza Vaccines.Influenza Other Respir Viruses. 2024 Mar;18(3):e13276. doi: 10.1111/irv.13276. Influenza Other Respir Viruses. 2024. PMID: 38513364 Free PMC article. Review.
Cited by
-
Reconstructed influenza A/H3N2 infection histories reveal variation in incidence and antibody dynamics over the life course.medRxiv [Preprint]. 2024 Apr 5:2024.03.18.24304371. doi: 10.1101/2024.03.18.24304371. medRxiv. 2024. Update in: PLoS Biol. 2024 Nov 7;22(11):e3002864. doi: 10.1371/journal.pbio.3002864. PMID: 38562868 Free PMC article. Updated. Preprint.
-
Avian Influenza A(H5N1) Neuraminidase Inhibition Antibodies in Healthy Adults after Exposure to Influenza A(H1N1)pdm09.Emerg Infect Dis. 2024 Jan;30(1):168-171. doi: 10.3201/eid3001.230756. Emerg Infect Dis. 2024. PMID: 38147510 Free PMC article.
-
Differential antigenic imprinting effects between influenza H1N1 hemagglutinin and neuraminidase in a mouse model.J Virol. 2025 Jan 31;99(1):e0169524. doi: 10.1128/jvi.01695-24. Epub 2024 Dec 5. J Virol. 2025. PMID: 39636110 Free PMC article.
-
Targeting neuraminidase: the next frontier for broadly protective influenza vaccines.Trends Immunol. 2024 Jan;45(1):11-19. doi: 10.1016/j.it.2023.11.001. Epub 2023 Dec 15. Trends Immunol. 2024. PMID: 38103991 Free PMC article. Review.
-
Reconstructed influenza A/H3N2 infection histories reveal variation in incidence and antibody dynamics over the life course.PLoS Biol. 2024 Nov 7;22(11):e3002864. doi: 10.1371/journal.pbio.3002864. eCollection 2024 Nov. PLoS Biol. 2024. PMID: 39509444 Free PMC article.
References
-
- Ng S, Nachbagauer R, Balmaseda A, Stadlbauer D, Ojeda S, Patel M, Rajabhathor A, Lopez R, Guglia AF, Sanchez N, Amanat F, Gresh L, Kuan G, Krammer F, Gordon A. 2019. Novel correlates of protection against pandemic H1N1 influenza A virus infection. Nat Med 25:962–967. doi:10.1038/s41591-019-0463-x. - DOI - PMC - PubMed
-
- Park J-K, Han A, Czajkowski L, Reed S, Athota R, Bristol T, Rosas LA, Cervantes-Medina A, Taubenberger JK, Memoli MJ. 2018. Evaluation of preexisting anti-hemagglutinin stalk antibody as a correlate of protection in a healthy volunteer challenge with influenza A/H1N1pdm Virus. mBio 9. doi:10.1128/mBio.02284-17. - DOI - PMC - PubMed
-
- Maier HE, Nachbagauer R, Kuan G, Ng S, Lopez R, Sanchez N, Stadlbauer D, Gresh L, Schiller A, Rajabhathor A, Ojeda S, Guglia AF, Amanat F, Balmaseda A, Krammer F, Gordon A. 2020. Pre-existing antineuraminidase antibodies are associated with shortened duration of influenza A(H1N1)pdm virus shedding and illness in naturally infected Adults. Clin Infect Dis 70:2290–2297. doi:10.1093/cid/ciz639. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
