

Hybrid breakdown in male reproduction between recently diverged Drosophila melanogaster populations has a complex and variable genetic architecture
- PMID: 37071601
- PMCID: PMC10309968
- DOI: 10.1093/evolut/qpad060
Hybrid breakdown in male reproduction between recently diverged Drosophila melanogaster populations has a complex and variable genetic architecture
Abstract
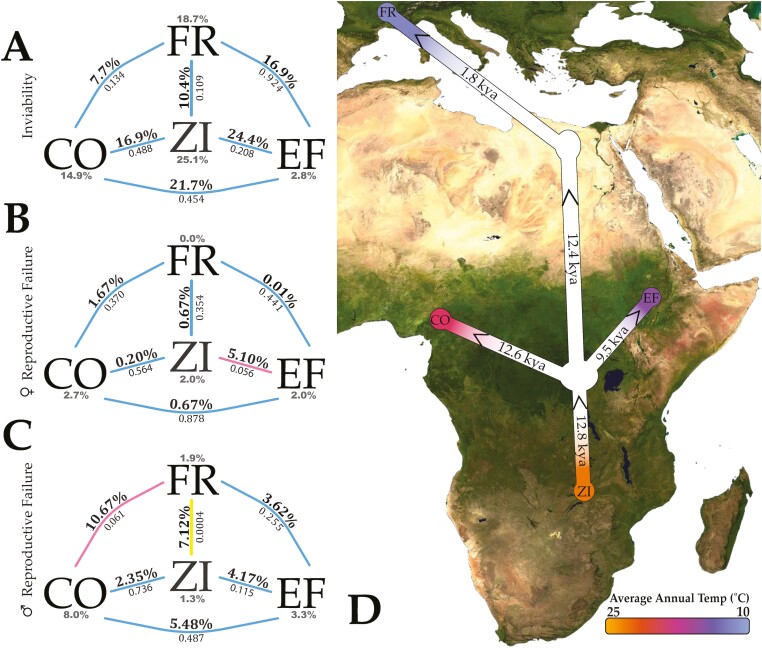
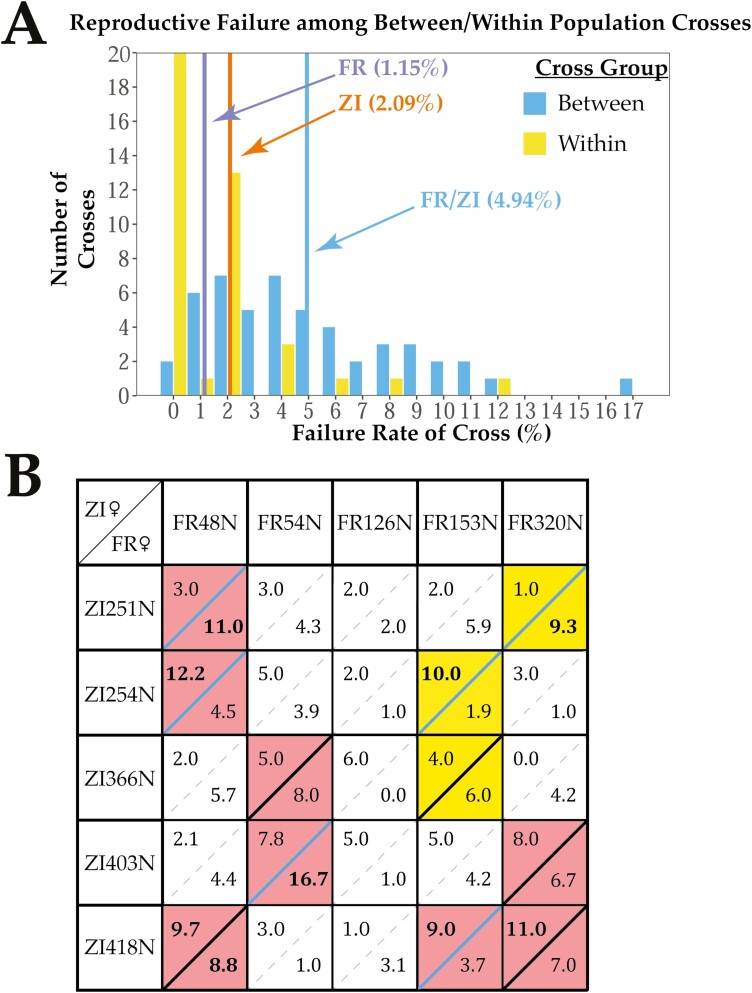
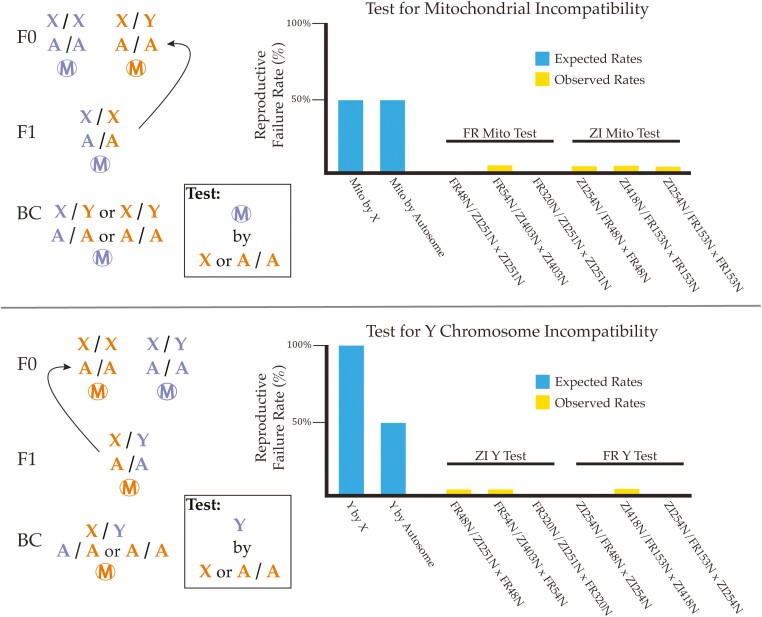
Secondary contact between formerly isolated populations may result in hybrid breakdown, in which untested allelic combinations in hybrids are maladaptive and limit genetic exchange. Studying early-stage reproductive isolation may yield key insights into the genetic architectures and evolutionary forces underlying the first steps toward speciation. Here, we leverage the recent worldwide expansion of Drosophila melanogaster to test for hybrid breakdown between populations that diverged within the last 13,000 years. We found clear evidence for hybrid breakdown in male reproduction, but not female reproduction or viability, supporting the prediction that hybrid breakdown affects the heterogametic sex first. The frequency of non-reproducing F2 males varied among different crosses involving the same southern African and European populations, as did the qualitative effect of cross direction, implying a genetically variable basis of hybrid breakdown and a role for uniparentally inherited factors. The levels of breakdown observed in F2 males were not recapitulated in backcrossed individuals, consistent with the existence of incompatibilities with at least three partners. Thus, some of the very first steps toward reproductive isolation could involve incompatibilities with complex and variable genetic architectures. Collectively, our findings emphasize this system's potential for future studies on the genetic and organismal basis of early-stage reproductive isolation.
Keywords: Drosophila melanogaster; genetic incompatibilities; genetic variation; hybrid breakdown; male reproduction; reproductive isolation.
© The Author(s) 2023. Published by Oxford University Press on behalf of The Society for the Study of Evolution (SSE).
Conflict of interest statement
Conflict of interest: The authors declare that no conflicts of interest exist.
Figures
References
-
- Alipaz, J. A., Karr, T. L., & Wu, C. I. (2005). Evolution of sexual isolation in laboratory populations: fitness differences between mating types and the associated hybrid incompatibilities. The American Naturalist, 165(4), 429–438. - PubMed
-
- Bateson, W. (1909) Heredity and variation in modern lights. Darwin and modern science: Essays in commemoration of the centenary of the birth of Charles Darwin and of the fiftieth anniversary of the publication of the origin of species. Cambridge University Press.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous