

Pigmentation and TYRP1 expression are mediated by zinc through the early secretory pathway-resident ZNT proteins
- PMID: 37072620
- PMCID: PMC10113262
- DOI: 10.1038/s42003-023-04640-5
Pigmentation and TYRP1 expression are mediated by zinc through the early secretory pathway-resident ZNT proteins
Abstract
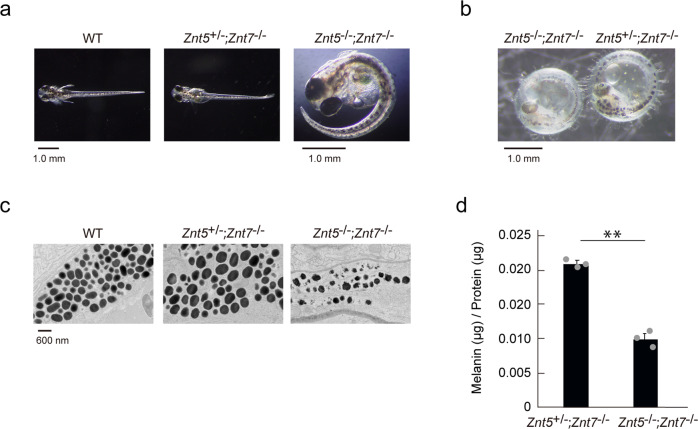
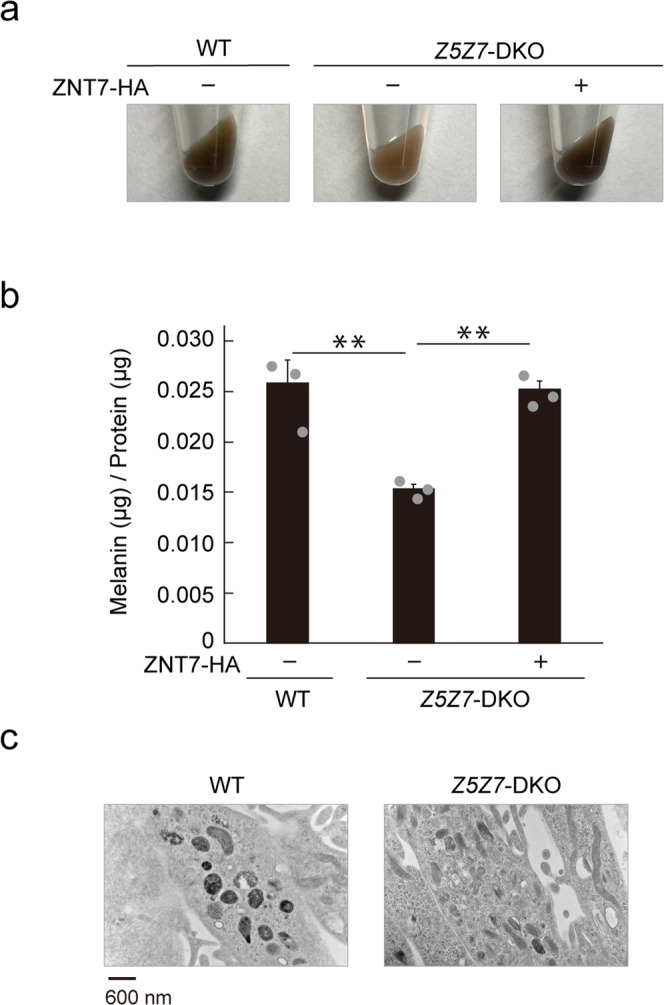
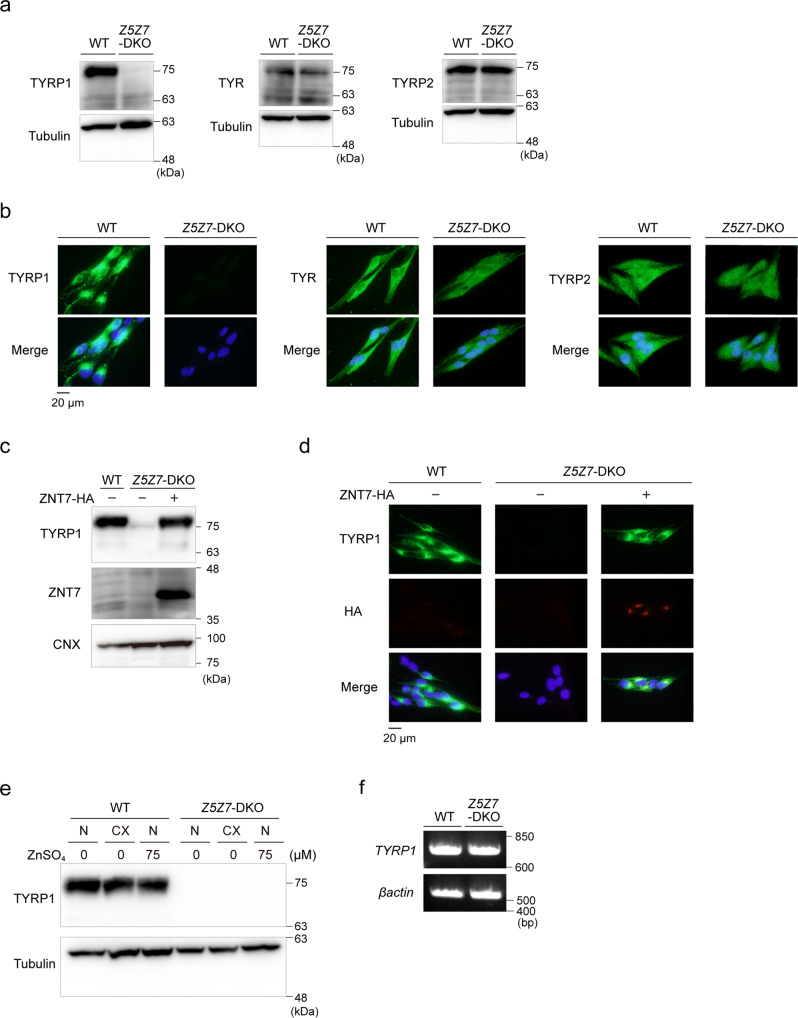
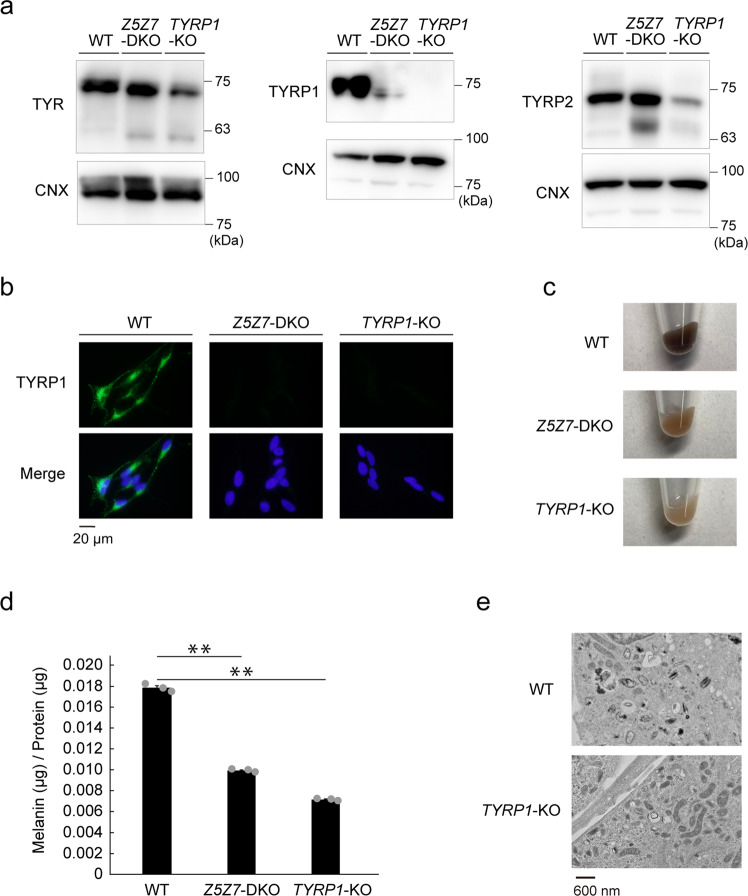
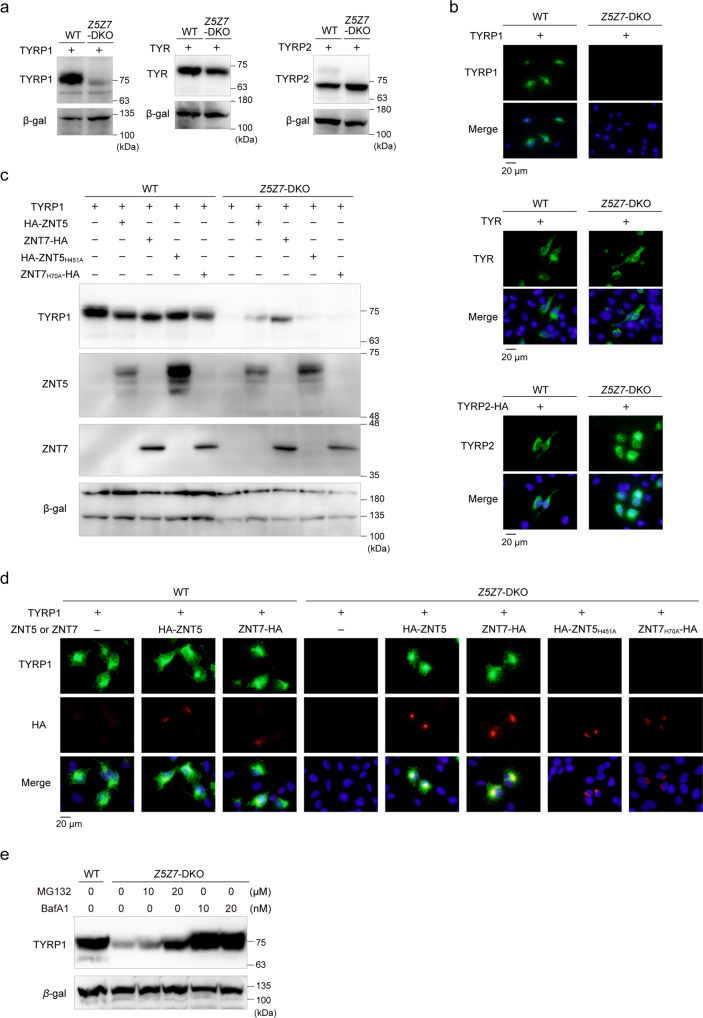
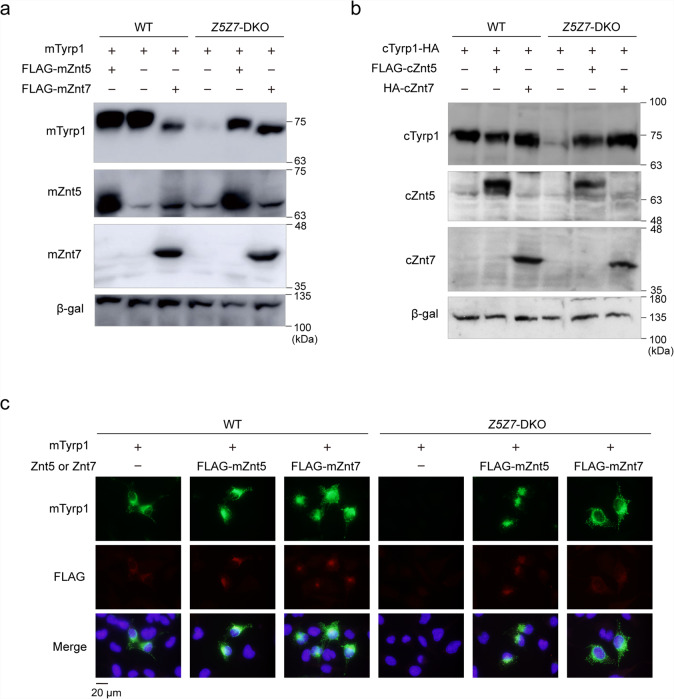
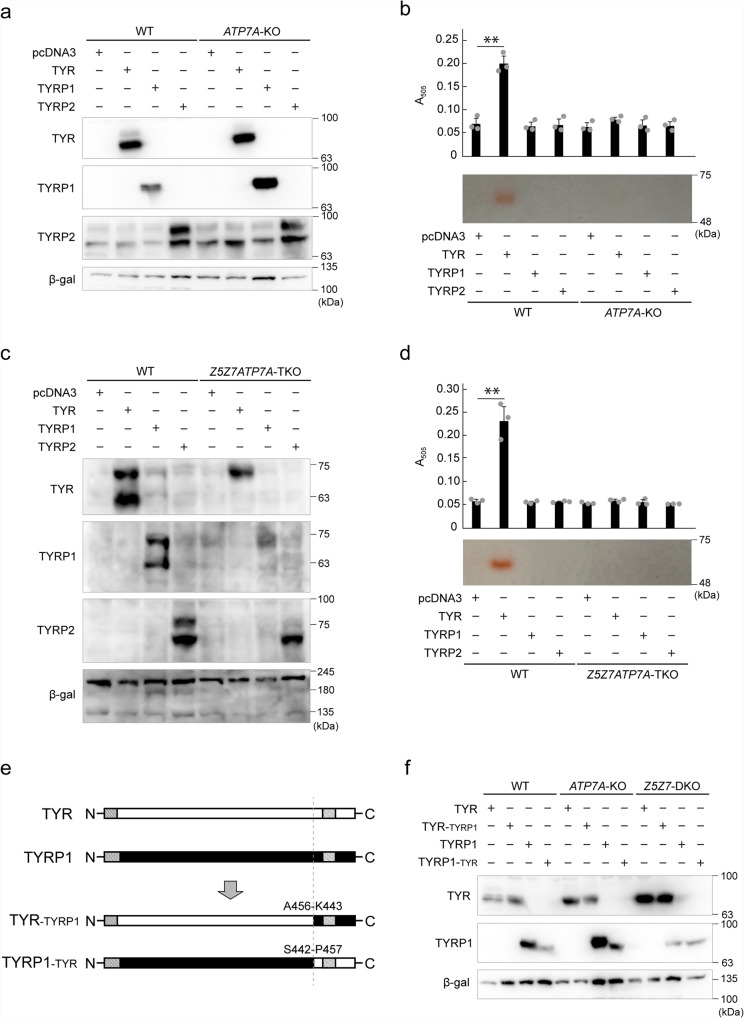
Tyrosinase (TYR) and tyrosinase-related proteins 1 and 2 (TYRP1 and TYRP2) are essential for pigmentation. They are generally classified as type-3 copper proteins, with binuclear copper active sites. Although there is experimental evidence for a copper cofactor in TYR, delivered via the copper transporter, ATP7A, the presence of copper in TYRP1 and TYRP2 has not been demonstrated. Here, we report that the expression and function of TYRP1 requires zinc, mediated by ZNT5-ZNT6 heterodimers (ZNT5-6) or ZNT7-ZNT7 homodimers (ZNT7). Loss of ZNT5-6 and ZNT7 function results in hypopigmentation in medaka fish and human melanoma cells, and is accompanied by immature melanosomes and reduced melanin content, as observed in TYRP1 dysfunction. The requirement of ZNT5-6 and ZNT7 for TYRP1 expression is conserved in human, mouse, and chicken orthologs. Our results provide novel insights into the pigmentation process and address questions regarding metalation in tyrosinase protein family.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Zinc transport via ZNT5-6 and ZNT7 is critical for cell surface glycosylphosphatidylinositol-anchored protein expression.J Biol Chem. 2022 Jun;298(6):102011. doi: 10.1016/j.jbc.2022.102011. Epub 2022 May 4. J Biol Chem. 2022. PMID: 35525268 Free PMC article.
-
Dissecting the Process of Activation of Cancer-promoting Zinc-requiring Ectoenzymes by Zinc Metalation Mediated by ZNT Transporters.J Biol Chem. 2017 Feb 10;292(6):2159-2173. doi: 10.1074/jbc.M116.763946. Epub 2016 Dec 27. J Biol Chem. 2017. PMID: 28028180 Free PMC article.
-
Detailed analyses of the crucial functions of Zn transporter proteins in alkaline phosphatase activation.J Biol Chem. 2020 Apr 24;295(17):5669-5684. doi: 10.1074/jbc.RA120.012610. Epub 2020 Mar 16. J Biol Chem. 2020. PMID: 32179649 Free PMC article.
-
Structure and Function of Human Tyrosinase and Tyrosinase-Related Proteins.Chemistry. 2018 Jan 2;24(1):47-55. doi: 10.1002/chem.201704410. Epub 2017 Nov 28. Chemistry. 2018. PMID: 29052256 Review.
-
Membrane transport proteins in melanosomes: Regulation of ions for pigmentation.Biochim Biophys Acta Biomembr. 2020 Dec 1;1862(12):183318. doi: 10.1016/j.bbamem.2020.183318. Epub 2020 Apr 22. Biochim Biophys Acta Biomembr. 2020. PMID: 32333855 Free PMC article. Review.
Cited by
-
Mammalian copper homeostasis: physiological roles and molecular mechanisms.Physiol Rev. 2025 Jan 1;105(1):441-491. doi: 10.1152/physrev.00011.2024. Epub 2024 Aug 22. Physiol Rev. 2025. PMID: 39172219 Free PMC article. Review.
-
A Splicing Mutation in mitfa is Involved in the Depigmentation of Cavefish Triplophysa rosa.Mol Biol Evol. 2025 Jul 30;42(8):msaf175. doi: 10.1093/molbev/msaf175. Mol Biol Evol. 2025. PMID: 40709436 Free PMC article.
-
Self-Delivering RNAi Compounds for Reduction of Hyperpigmentation.Clin Cosmet Investig Dermatol. 2024 Dec 27;17:3033-3044. doi: 10.2147/CCID.S498987. eCollection 2024. Clin Cosmet Investig Dermatol. 2024. PMID: 39741580 Free PMC article.
-
Copper in melanoma: At the crossroad of protumorigenic and anticancer roles.Redox Biol. 2025 Apr;81:103552. doi: 10.1016/j.redox.2025.103552. Epub 2025 Feb 15. Redox Biol. 2025. PMID: 39970778 Free PMC article. Review.
-
In-depth transcriptomic analysis of Anopheles gambiae hemocytes uncovers novel genes and the oenocytoid developmental lineage.BMC Genomics. 2024 Jan 19;25(1):80. doi: 10.1186/s12864-024-09986-6. BMC Genomics. 2024. PMID: 38243165 Free PMC article.
