

Degradation of NLRP3 by p62-dependent-autophagy improves cognitive function in Alzheimer's disease by maintaining the phagocytic function of microglia
- PMID: 37072933
- PMCID: PMC10493665
- DOI: 10.1111/cns.14219
Degradation of NLRP3 by p62-dependent-autophagy improves cognitive function in Alzheimer's disease by maintaining the phagocytic function of microglia
Abstract
Background: Activation of the NLRP3 inflammasome promotes microglia to secrete inflammatory cytokines and induce pyroptosis, leading to impaired phagocytic and clearance functions of microglia in Alzheimer's disease (AD). This study found that the autophagy-associated protein p62 interacts with NLRP3, which is the rate-limiting protein of the NLRP3 inflammasome. Thus, we aimed to prove that the degradation of NLRP3 occurs through the autophagy-lysosome pathway (ALP) and also demonstrate its effects on the function of microglia and pathological changes in AD.
Methods: The 5XFAD/NLRP3-KO mouse model was established to study the effect of NLRP3 reduction on AD. Behavioral experiments were conducted to assess the cognitive function of the mice. In addition, immunohistochemistry was used to evaluate the deposition of Aβ plaques and morphological changes in microglia. BV2 cells treated with lipopolysaccharide (LPS) followed by Aβ1-42 oligomers were used as in vitro AD inflammation models and transfected with lentivirus to regulate the expression of the target protein. The pro-inflammatory status and function of BV2 cells were detected by flow cytometry and immunofluorescence (IF). Co-immunoprecipitation, mass spectrometry, IF, Western blot (WB), quantitative real-time PCR, and RNA-seq analysis were used to elucidate the mechanisms of molecular regulation.
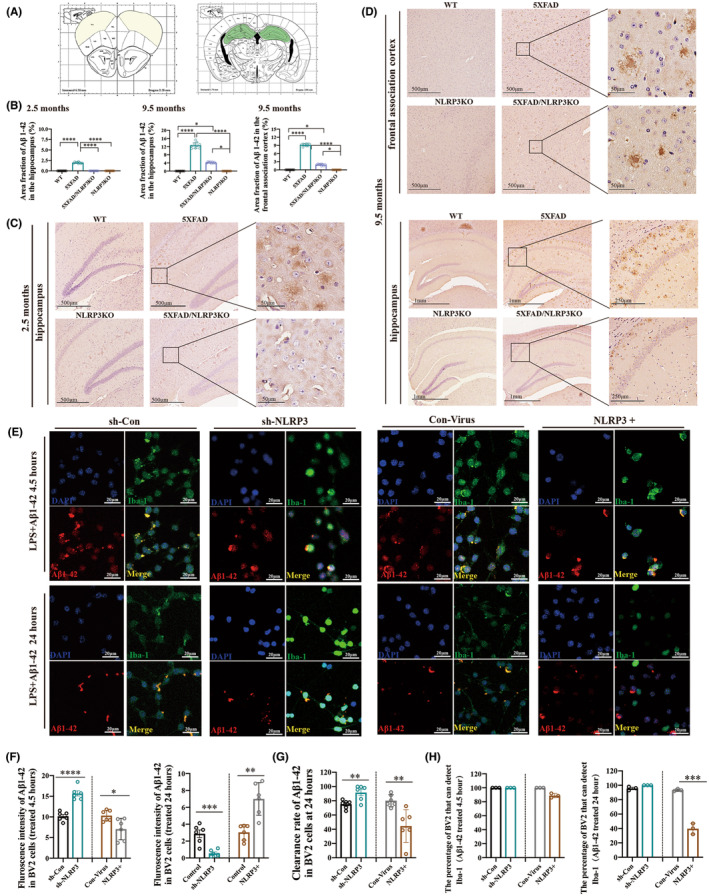
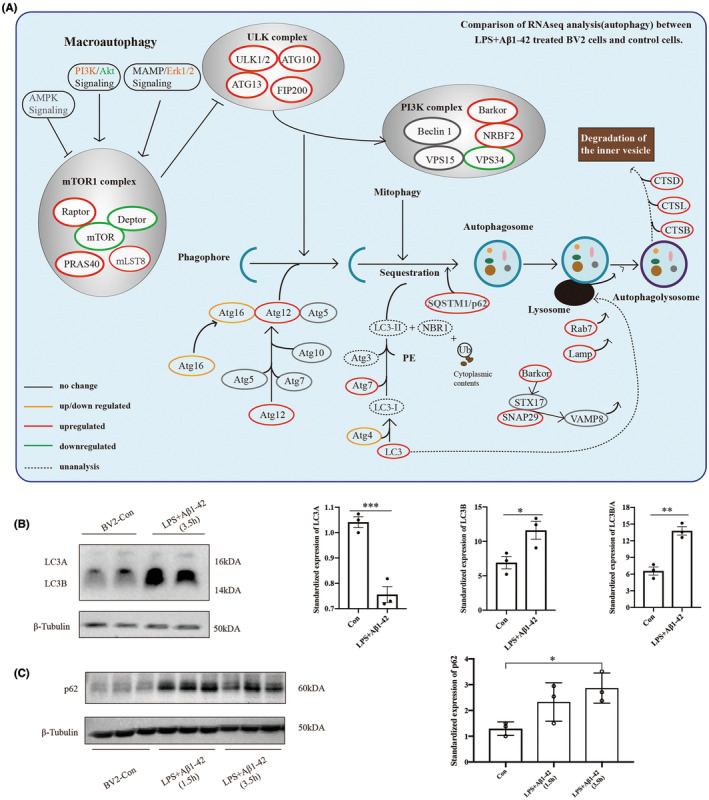
Results: Cognitive function was improved in the 5XFAD/NLRP3-KO mouse model by reducing the pro-inflammatory response of microglia and maintaining the phagocytic and clearance function of microglia to the deposited Aβ plaque. The pro-inflammatory function and pyroptosis of microglia were regulated by NLRP3 expression. Ubiquitinated NLRP3 can be recognized by p62 and degraded by ALP, slowing down the proinflammatory function and pyroptosis of microglia. The expression of autophagy pathway-related proteins such as LC3B/A, p62 was increased in the AD model in vitro.
Conclusions: P62 recognizes and binds to ubiquitin-modified NLRP3. It plays a vital role in regulating the inflammatory response by participating in ALP-associated NLRP3 protein degradation, which improves cognitive function in AD by reducing the pro-inflammatory status and pyroptosis of microglia, thus maintaining its phagocytic function.
Keywords: Alzheimer's disease; NLRP3 inflammasome; autophagy; pyroptosis; sqstm1/p62; ubiquitin.
© 2023 The Authors. CNS Neuroscience & Therapeutics published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors have no relevant financial or nonfinancial interests to disclose.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
