

doi: 10.1038/s43586-022-00130-w.
Epub 2022 May 19.
In vivo hypermutation and continuous evolution
Affiliations
- PMID: 37073402
- PMCID: PMC10108624
- DOI: 10.1038/s43586-022-00130-w
Item in Clipboard
In vivo hypermutation and continuous evolution
Nat Rev Methods Primers.
2022.
No abstract available
Keywords: Cheater mutations; Clonal interference; Consensus sequencing; Directed evolution; Greedy mutations; Hypermutation; Integration cassette; Processivity; Sign epistasis; Unique molecular identifier.
Conflict of interest statement
Competing interests The authors declare no competing interests.
Figures
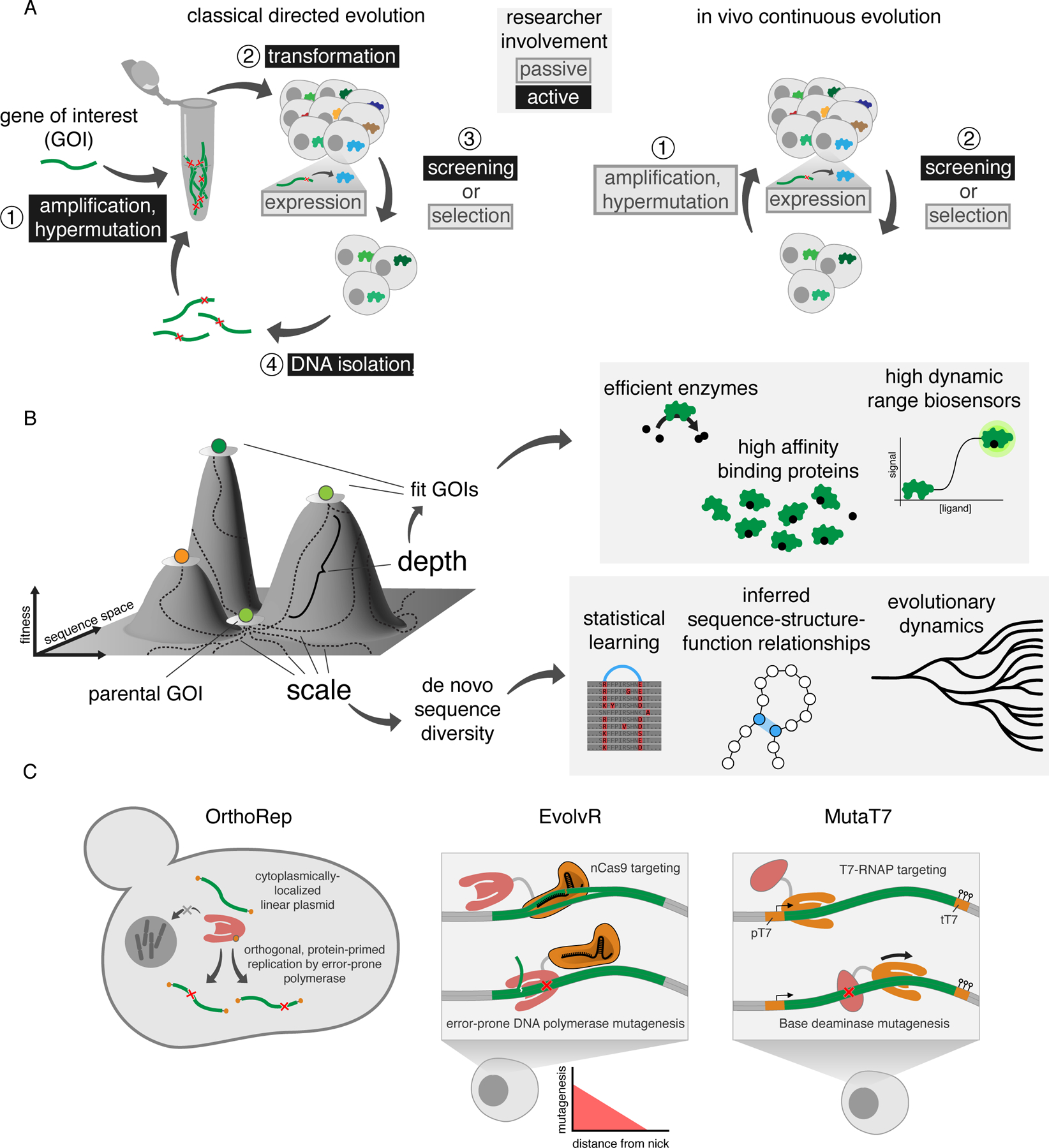
A) Comparison of classical directed evolution and in vivo continuous evolution. Classical directed evolution relies upon discrete steps performed in vitro (for example, hypermutation through error-prone PCR) and in vivo. Transformation, DNA isolation and PCR-based amplification and hypermutation are required to complete classical directed evolution cycles. In contrast, in vivo continuous evolution cycles can be carried out autonomously as cells grow. While amplification and hypermutation require cell culturing and therefore some researcher intervention, this dilution task is trivial enough to be considered passive. B) Depth and scale in continuous evolution enable broad applications. C) Depictions of OrthoRep, EvolvR and MutaT7. Each system achieves targeted hypermutation. OrthoRep and EvolvR both use error-prone DNA replication-based mutagenesis, with the distinction that in EvolvR, error-prone replication is not essential for propagation of the GOI since error-prone replication occurs in addition to replication by host DNA polymerases. MutaT7 utilizes base modification by a nucleotide base deaminase for mutagenesis. All three systems utilize distinct targeting methods. OrthoRep has only been applied in yeast, while both EvolvR and MutaT7 have been used in multiple model organisms. The OrthoRep accessory plasmid p2 is not depicted for simplicity. Note that termination of transcription and hypermutation by MutaT7 systems may be accomplished by terminators (depicted) or dCas9 (not depicted). pT7, T7 RNA polymerase promoter; tT7, T7 RNA polymerase terminator array.

A) a GOI is chosen for its predicted potential to evolve a desired activity. Often, the ability to carry out the desired activity at a low level is a promising starting point, although activities that are similar to the desired activity may be suitable as well. A host cell and a hypermutation system are also chosen at this stage. B) the researcher must design, build and test a selection or screen that is capable of enriching cells encoding GOIs with the desired function. This requires some cell-based mechanism of converting the activity of the GOI into a growth or optical signal, here symbolized as falling dominos. If available, GOI variants with known propensity for the desired activity, or ‘fitness’, may be used to test that the screen or selection is effective. C) the GOI and necessary components of the hypermutation system are encoded in the host cell. D) continuous evolution strategies are applied to evolve the desired activities. Different strategies offer different advantages and disadvantages and should be considered against each other. Labor, resources, feasible population size, durability and other factors can vary depending on the chosen strategy. E) following an evolution experiment that appears successful, GOIs contained in evolved populations should be isolated and characterized via sequencing and functional assays. The chosen approach will depend on the specific demands of the experiment and application.

A) Types of nucleotide substitutions. B) and C) Following mutation by DNA replication error (B) or deamination (C), the nucleotide at the corresponding position on the opposite DNA strand is not yet altered. This mismatch will result in a mutation if it is used as a template in a subsequent DNA replication event. Alternatively, the mismatch repair machinery of the cell may correctly repair this mismatch, preventing establishment of the mutation.
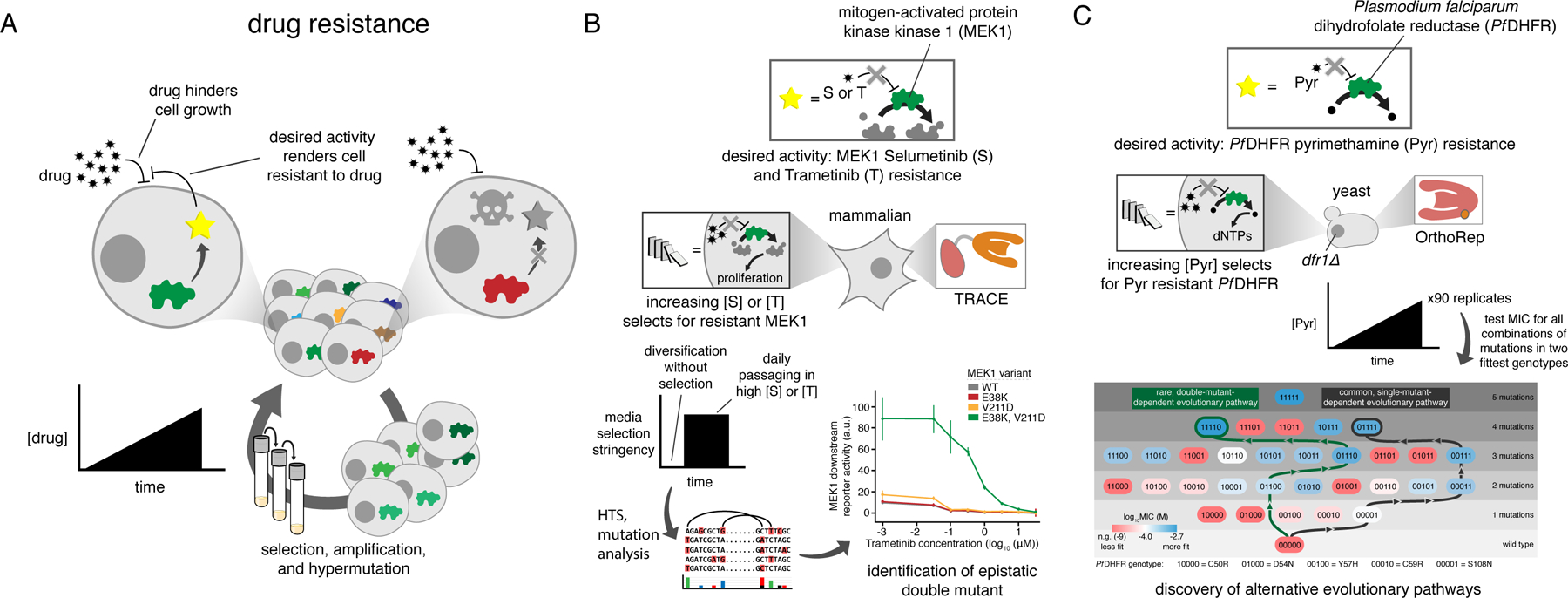
A) Generalized schematic for drug resistance continuous evolution experiments. A cycle with only one step is shown but is meant to represent continuous growth following desired selection schedules, for example serial passaging into media with increasing drug concentration. B) Continuous evolution of MEK1 with the TRACE MutaT7 system. The native activity of MEK1 is essential for proliferation in the mammalian cell line that was used (HEK293T) and is therefore drug selectable. MEK1 was targeted for mutagenesis and grown for a short initial period without selection for drug resistance to generate mutational diversity. Selection was then applied using a static high concentration of either of the two drugs being studied. C) Continuous evolution of DHFR to study drug resistance using OrthoRep. DHFR activity is essential for nucleotide biosynthesis and is therefore selectable in cells deleted for the native enzyme. Ninety replicate cultures were passaged in parallel with gradually increasing drug concentration, and Sanger sequencing of populations revealed two genotypes that commonly become fixed in populations. All combinations of the mutations in these two genotypes were individually cloned and tested in isolation to determine fitness and understand the accessibility of different mutational pathways leading to different outcomes. MIC, minimum inhibitory concentration.
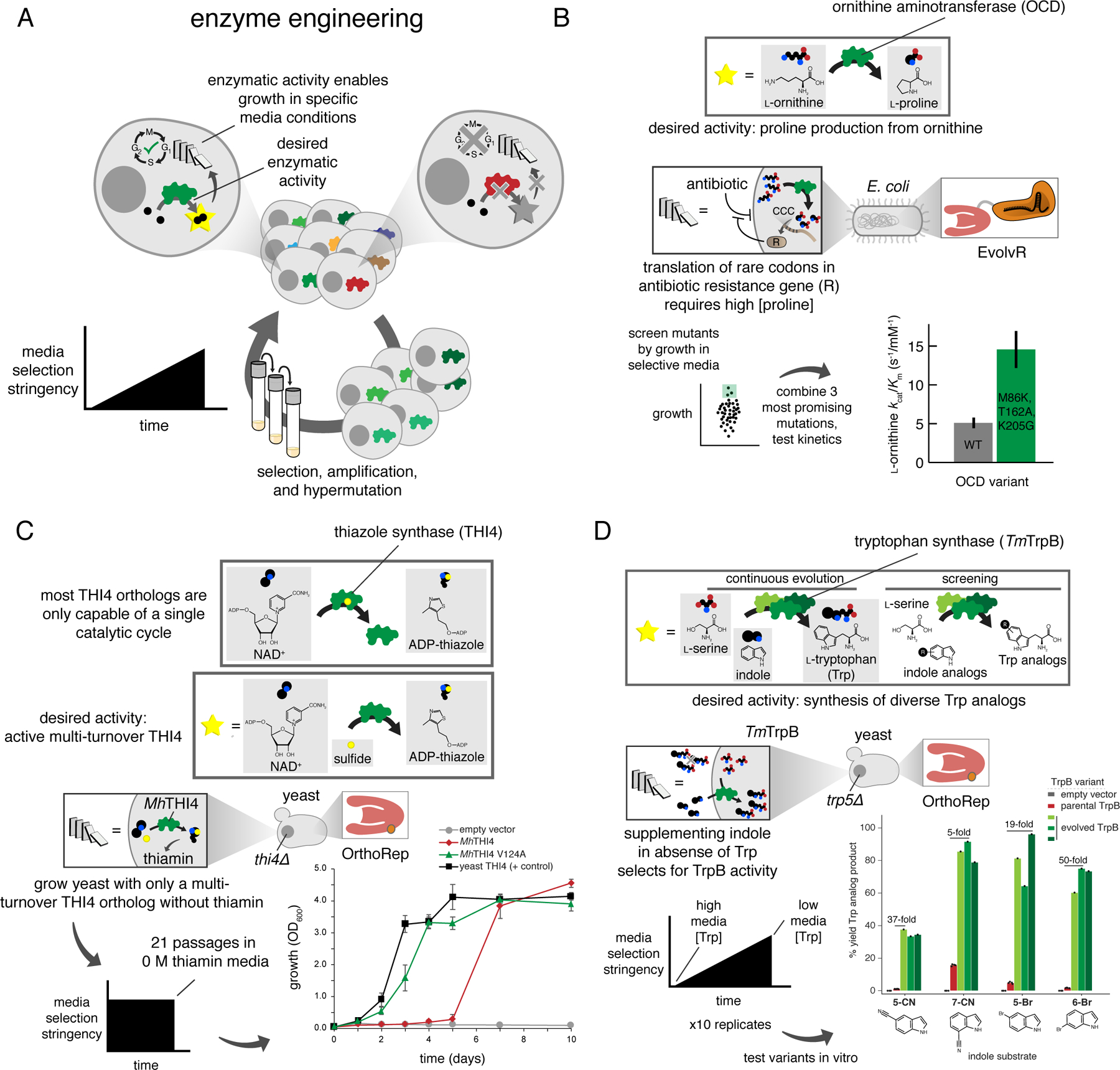
A) Generalized schematic for enzyme engineering through continuous evolution. A cycle with only one step is shown, as experiments are typically carried out using growth-based selections. This means all of the necessary steps for evolution are accomplished by the cell concurrently when provided with a consistent supply of media with increasing selection stringency. Media selection stringency modification is typically accomplished through reducing the concentration of a required nutrient whose production is made to be dependent upon GOI activity. B) Continuous evolution of OCD for improved proline production. Following diversification with EvolvR, clones underwent a single round of screening on the basis of colony size prior to testing for growth. C) Adaptation of a multi-turnover thiazole synthase from the prokaryote Mucinivorans hirudinis for activity in the plant-like cellular environment of yeast. D) Continuous evolution of tryptophan synthase from the thermophile Thermotoga maritima (TmTrpB) to generate enzymes with diverse substrate promiscuities.
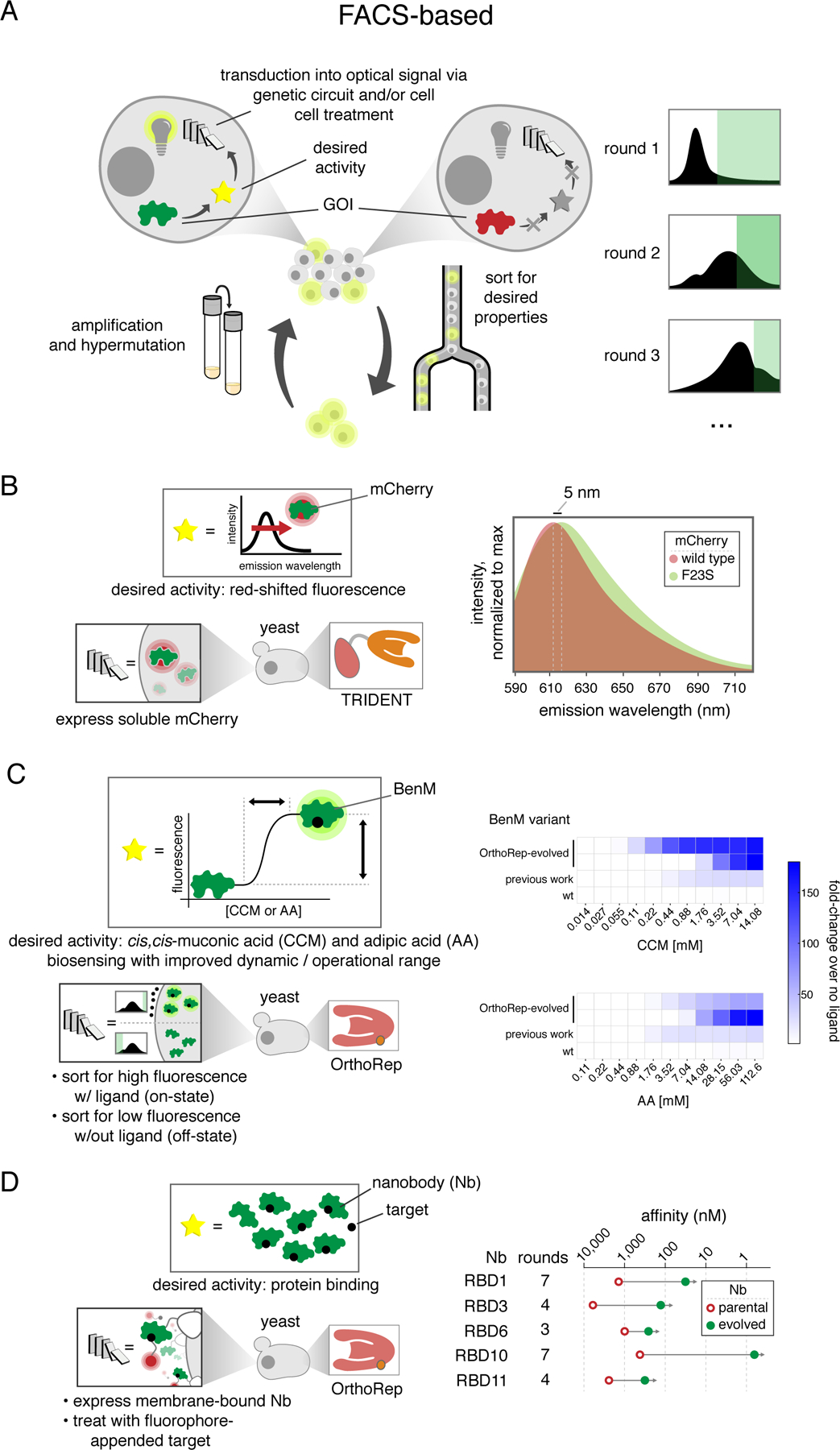
A) Generalized schematic for continuous evolution experiments that employ FACS-based high throughput screening to evolve GOIs with a desired activity. Unlike with selections, screening and amplification must be carried out in two discrete steps. B) Evolution of a red-shifted mCherry using the TRIDENT MutaT7 system. C) Evolution of the cis,cis-muconic acid biosensor BenM to exhibit higher dynamic range and broader operational range. D) Nanobody evolution using OrthoRep for diversification and yeast display for FACS-based screening.
References
-
- Arnold FH Design by Directed Evolution. Acc. Chem. Res 31, 125–131 (1998).
-
- Packer MS & Liu DR Methods for the directed evolution of proteins. Nat. Rev. Genet 16, 379–394 (2015). - PubMed
-
- Chen K & Arnold FH Engineering new catalytic activities in enzymes. Nat. Catal 3, 203–213 (2020).
Grants and funding
LinkOut - more resources
Full Text Sources