

The Impact of Mmu17 Non-Hsa21 Orthologous Genes in the Ts65Dn Mouse Model of Down Syndrome: The Gold Standard Refuted
- PMID: 37074246
- PMCID: PMC10330375
- DOI: 10.1016/j.biopsych.2023.02.012
The Impact of Mmu17 Non-Hsa21 Orthologous Genes in the Ts65Dn Mouse Model of Down Syndrome: The Gold Standard Refuted
Abstract
Background: Despite successful preclinical treatment studies to improve neurocognition in the Ts65Dn mouse model of Down syndrome, translation to humans has failed. This raises questions about the appropriateness of the Ts65Dn mouse as the gold standard. We used the novel Ts66Yah mouse that carries an extra chromosome and the identical segmental Mmu16 trisomy as Ts65Dn without the Mmu17 non-Hsa21 orthologous region.
Methods: Forebrains from embryonic day 18.5 Ts66Yah and Ts65Dn mice, along with euploid littermate controls, were used for gene expression and pathway analyses. Behavioral experiments were performed in neonatal and adult mice. Because male Ts66Yah mice are fertile, parent-of-origin transmission of the extra chromosome was studied.
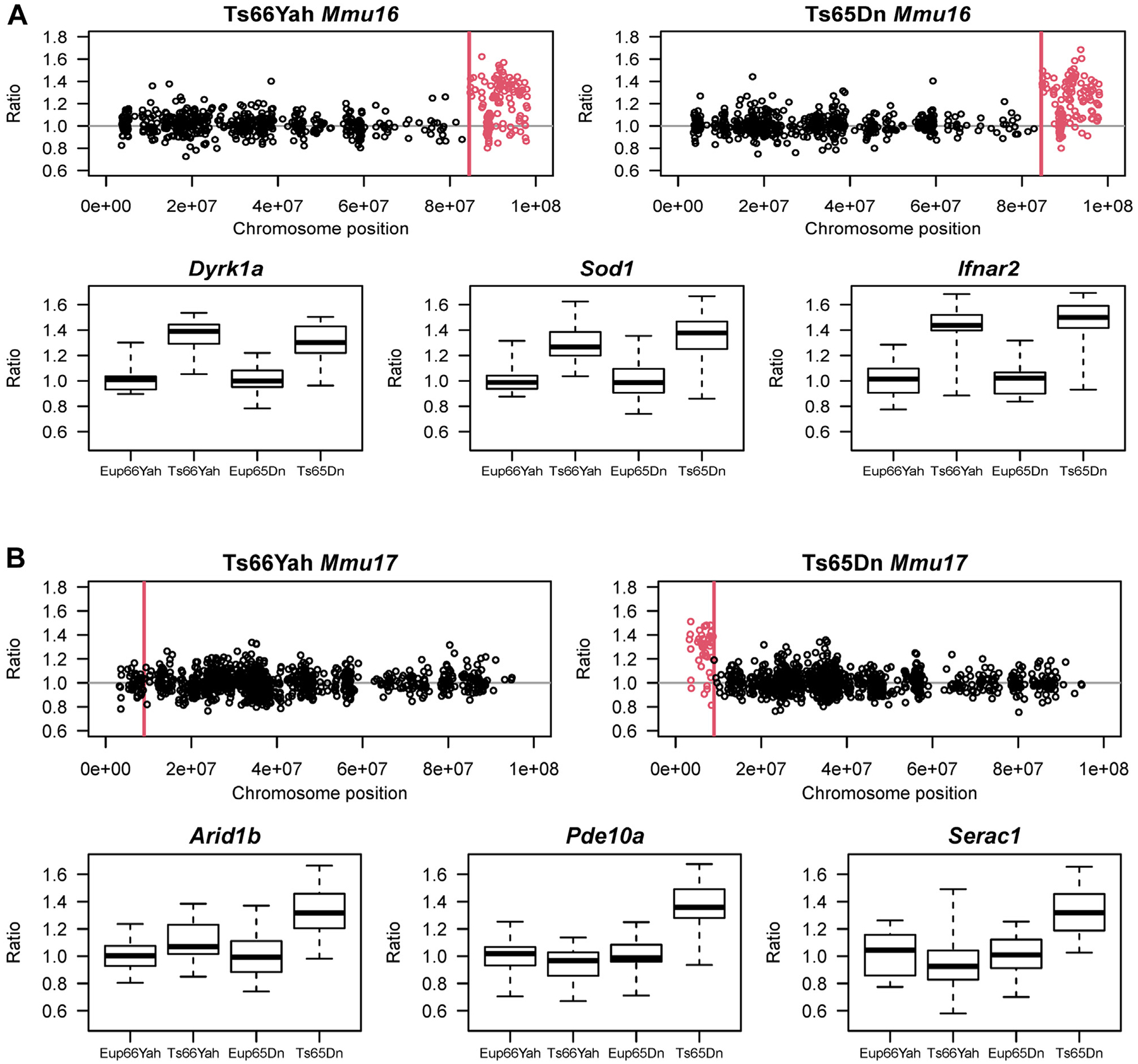
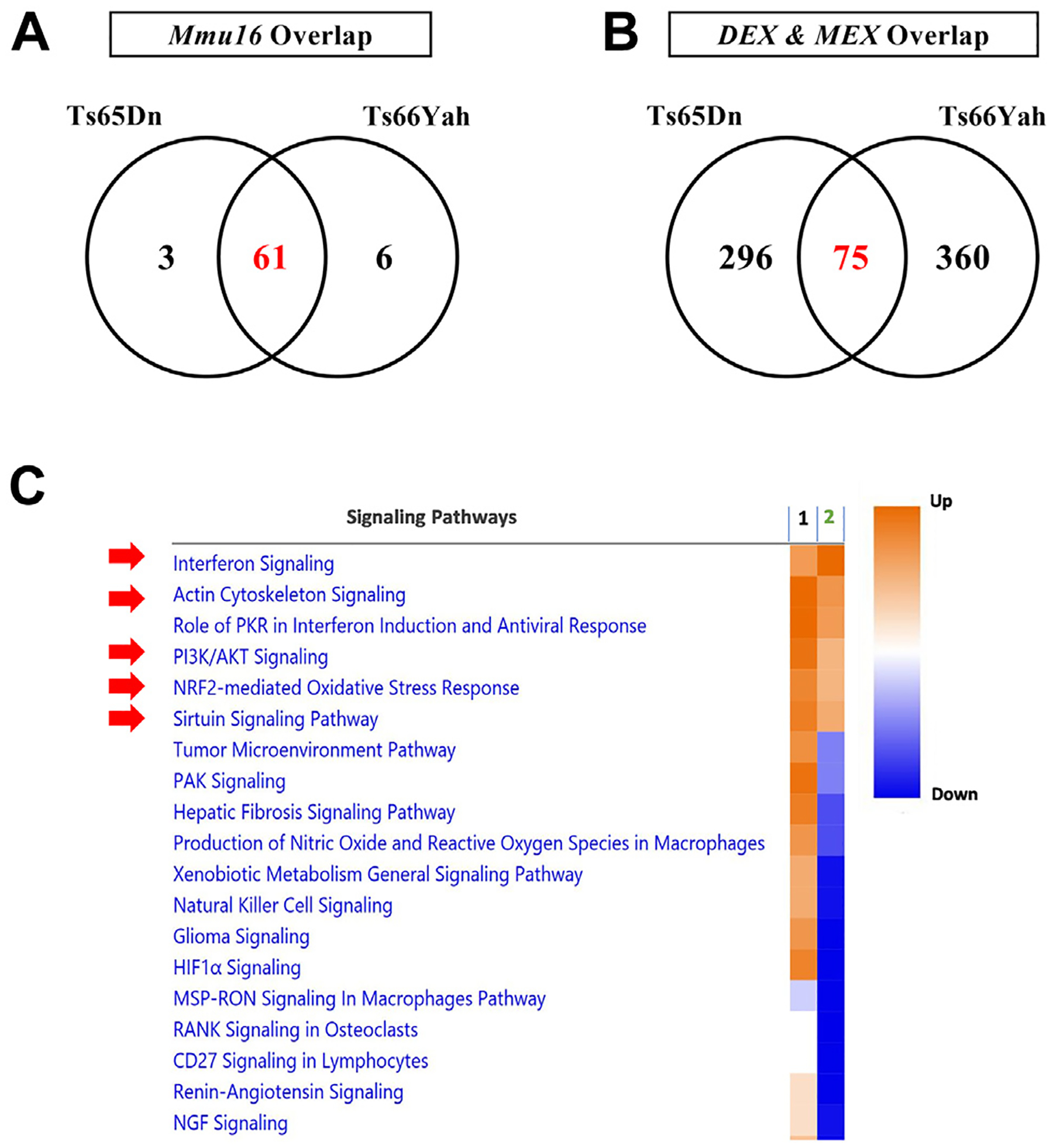
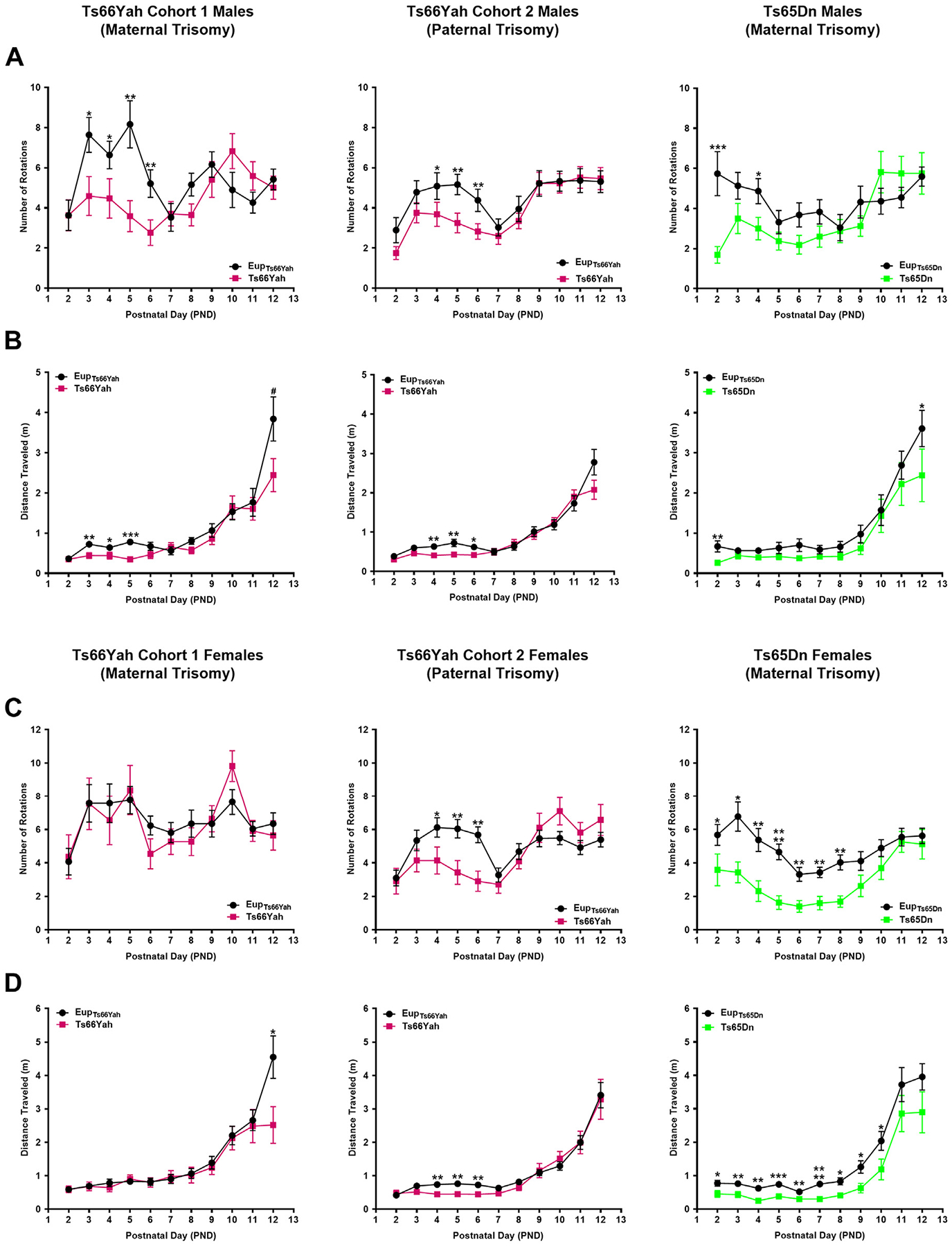
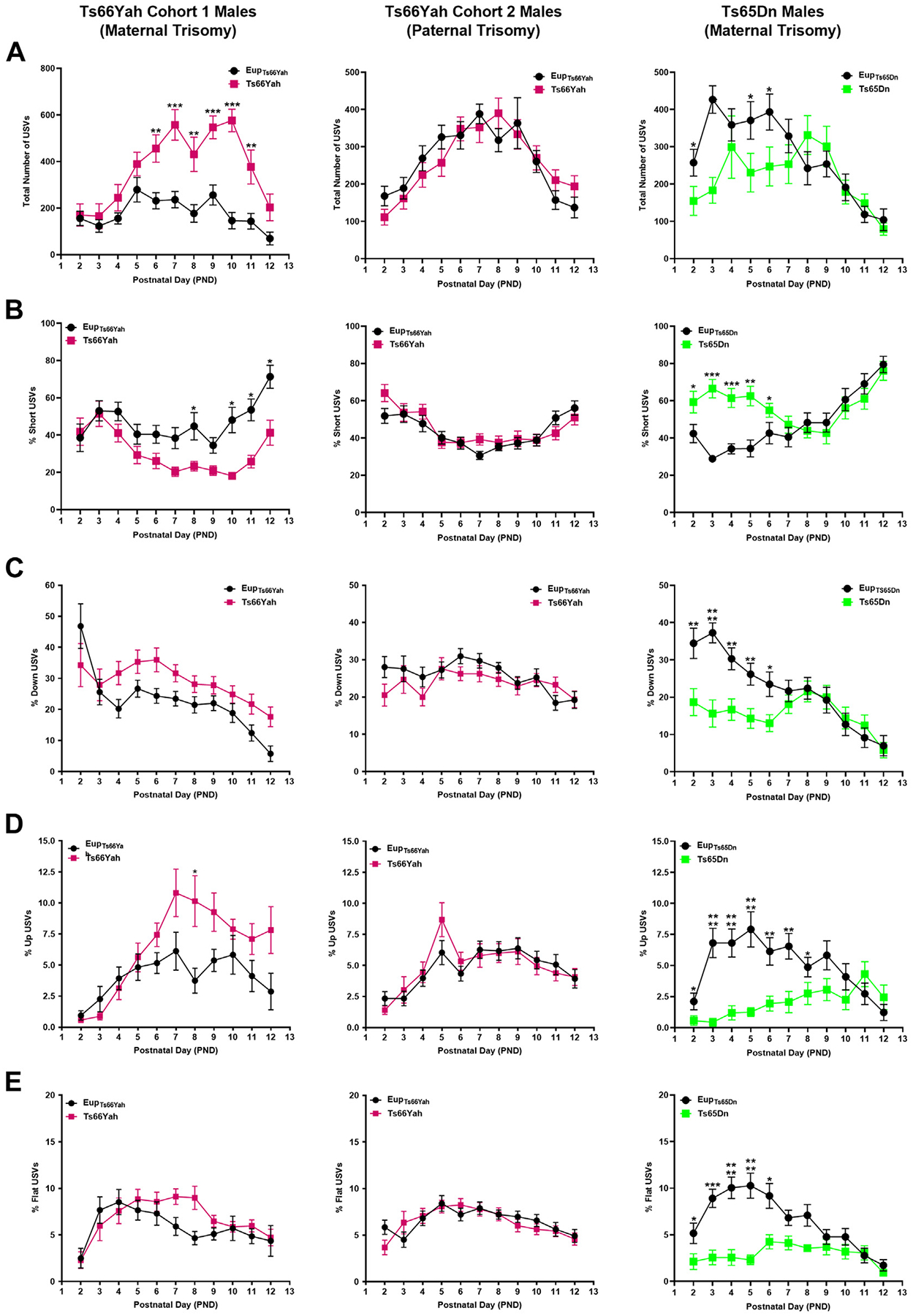
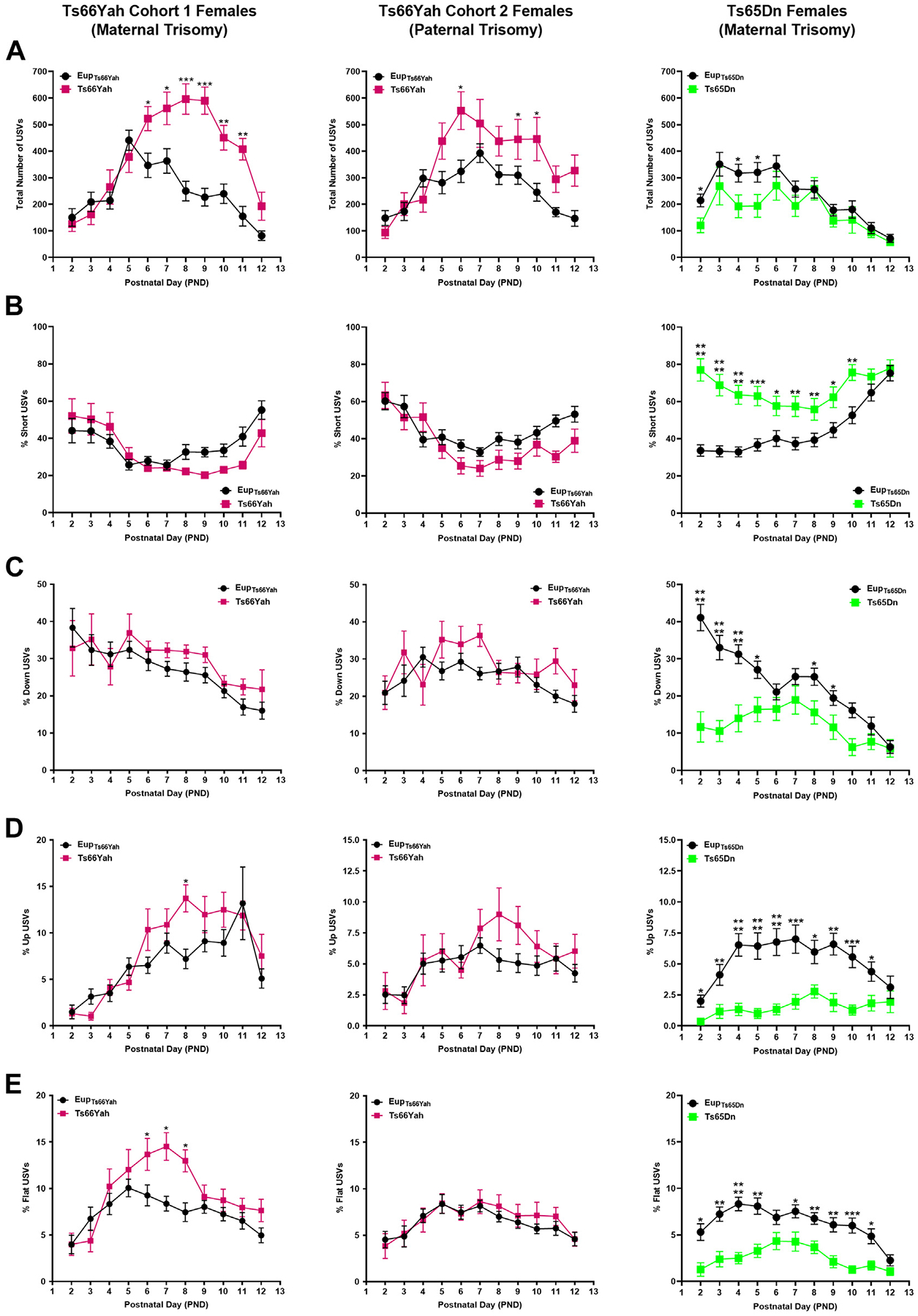
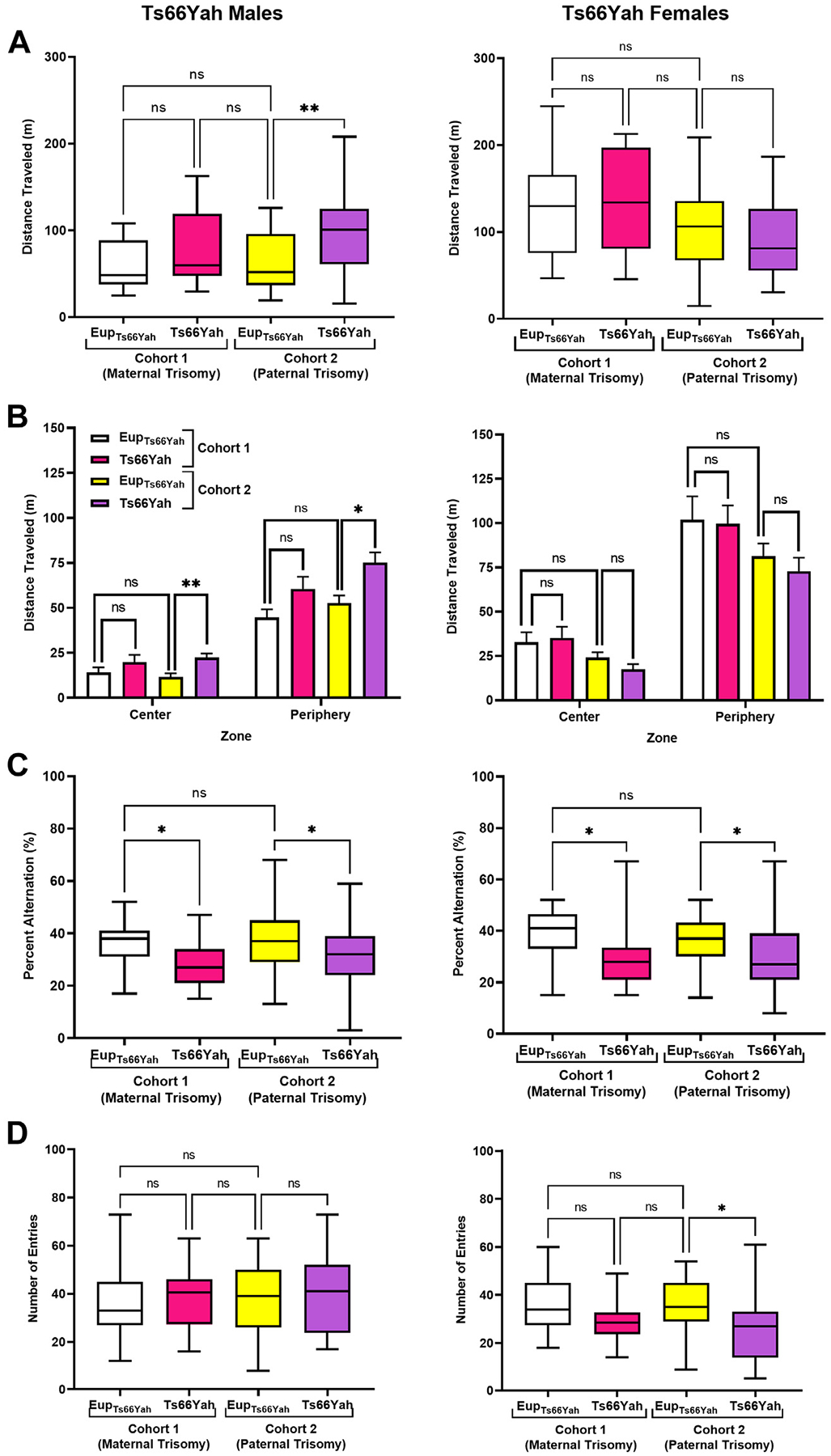
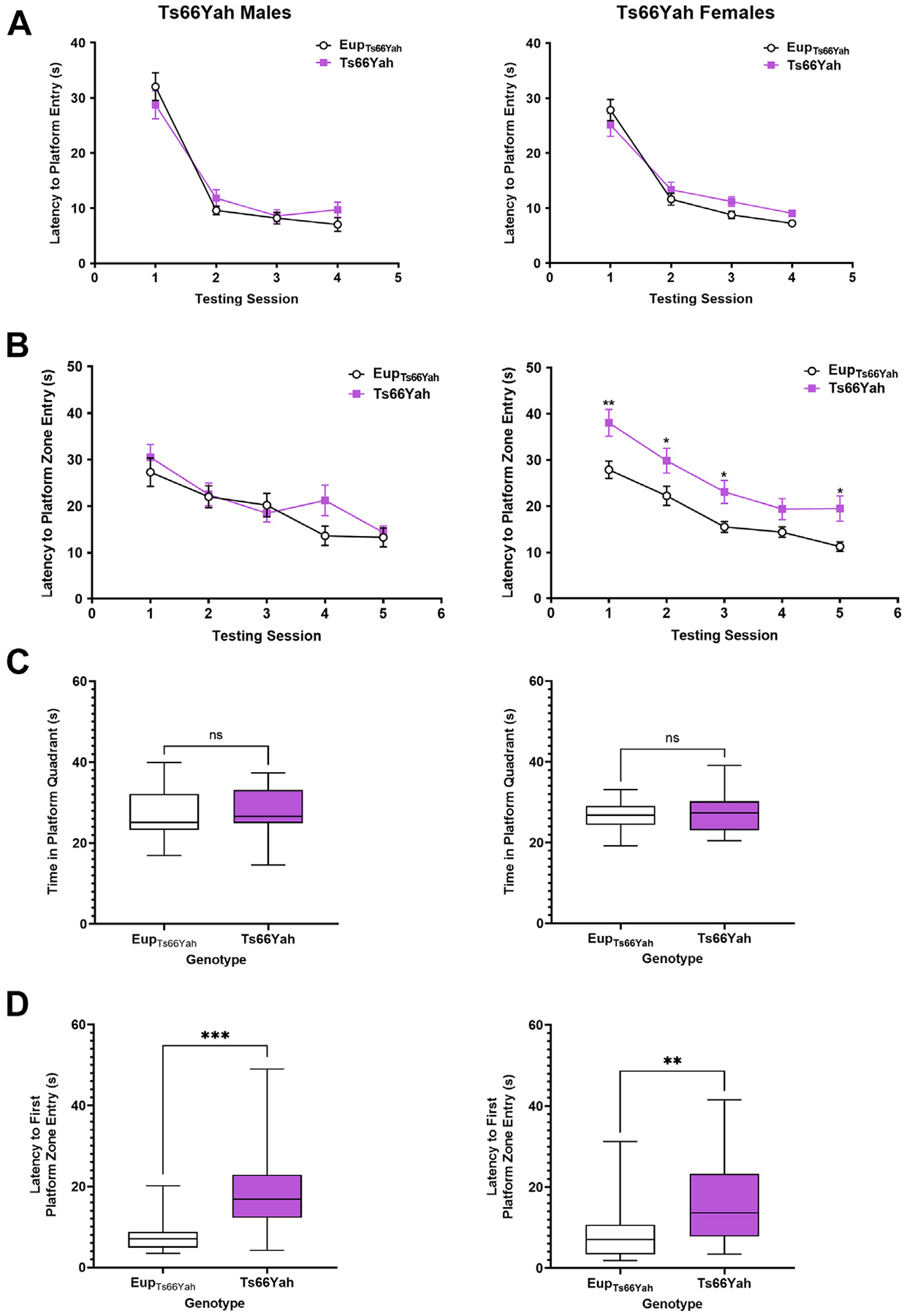
Results: Forty-five protein-coding genes mapped to the Ts65Dn Mmu17 non-Hsa21 orthologous region; 71%-82% are expressed during forebrain development. Several of these genes are uniquely overexpressed in Ts65Dn embryonic forebrain, producing major differences in dysregulated genes and pathways. Despite these differences, the primary Mmu16 trisomic effects were highly conserved in both models, resulting in commonly dysregulated disomic genes and pathways. Delays in motor development, communication, and olfactory spatial memory were present in Ts66Yah but more pronounced in Ts65Dn neonates. Adult Ts66Yah mice showed milder working memory deficits and sex-specific effects in exploratory behavior and spatial hippocampal memory, while long-term memory was preserved.
Conclusions: Our findings suggest that triplication of the non-Hsa21 orthologous Mmu17 genes significantly contributes to the phenotype of the Ts65Dn mouse and may explain why preclinical trials that used this model have unsuccessfully translated to human therapies.
Keywords: Down syndrome; Mmu17 non-Hsa21 orthologous genes; Mouse models; Phenotype; Trisomy 21; Ts65Dn; Ts66Yah.
Published by Elsevier Inc.
Figures
References
-
- Davisson MT, Schmidt C, Akeson EC (1990): Segmental trisomy of murine chromosome 16: A new model system for studying Down syndrome. Prog Clin Biol Res 360:263–280. - PubMed
-
- Davisson MT, Schmidt C, Reeves RH, Irving NG, Akeson EC, Harris BS, Bronson RT (1993): Segmental trisomy as a mouse model for Down syndrome. Prog Clin Biol Res 384:117–133. - PubMed
-
- Muñiz Moreno MDM, Brault V, Birling MC, Pavlovic G, Herault Y (2020): Modeling Down syndrome in animals from the early stage to the 4.0 models and next. Prog Brain Res 251:91–143. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
