

New High-Throughput Screen Discovers Novel Ligands of Full-Length Nuclear Receptor LRH-1
- PMID: 37074920
- PMCID: PMC10404069
- DOI: 10.1021/acschembio.2c00805
New High-Throughput Screen Discovers Novel Ligands of Full-Length Nuclear Receptor LRH-1
Abstract
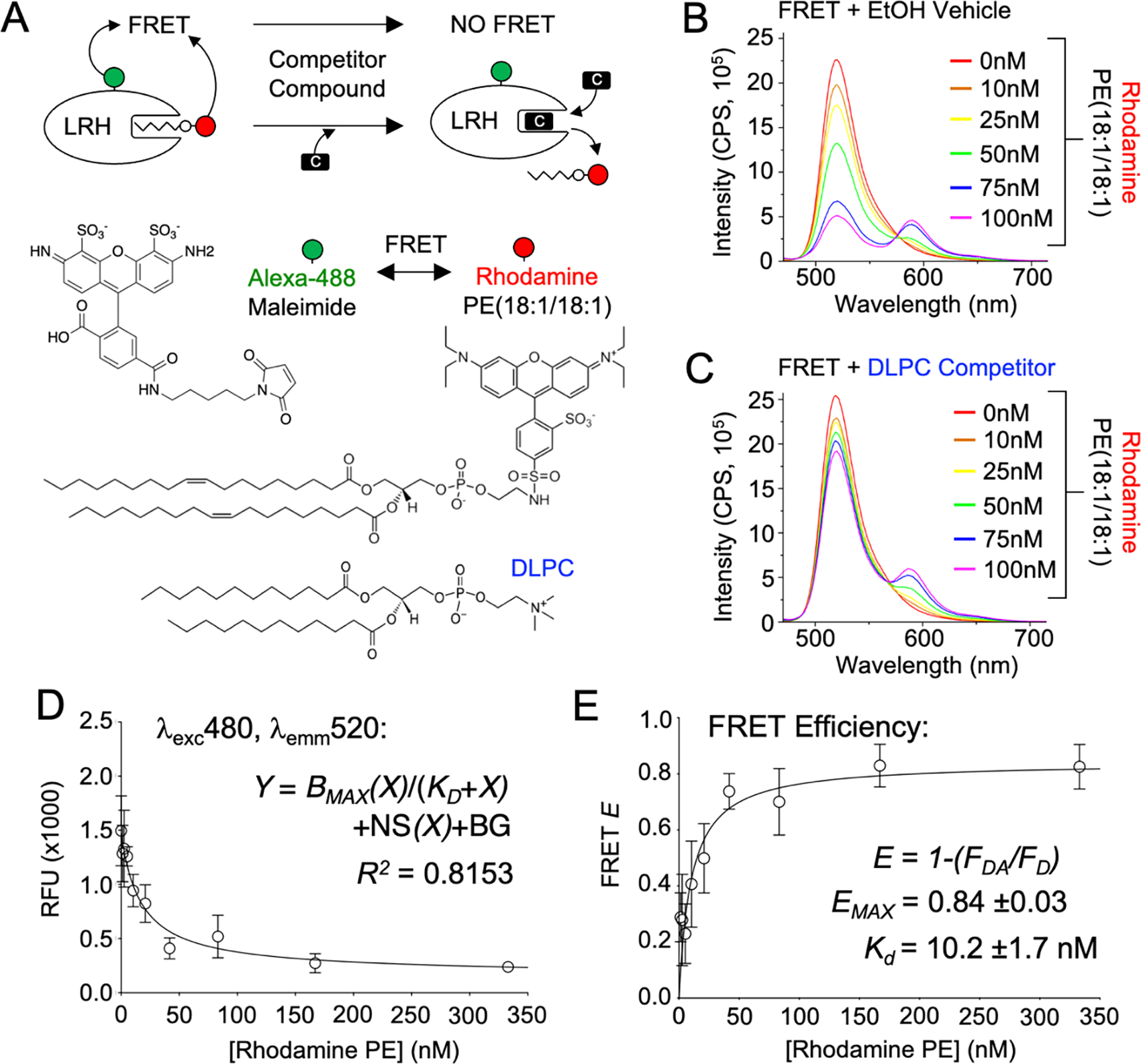
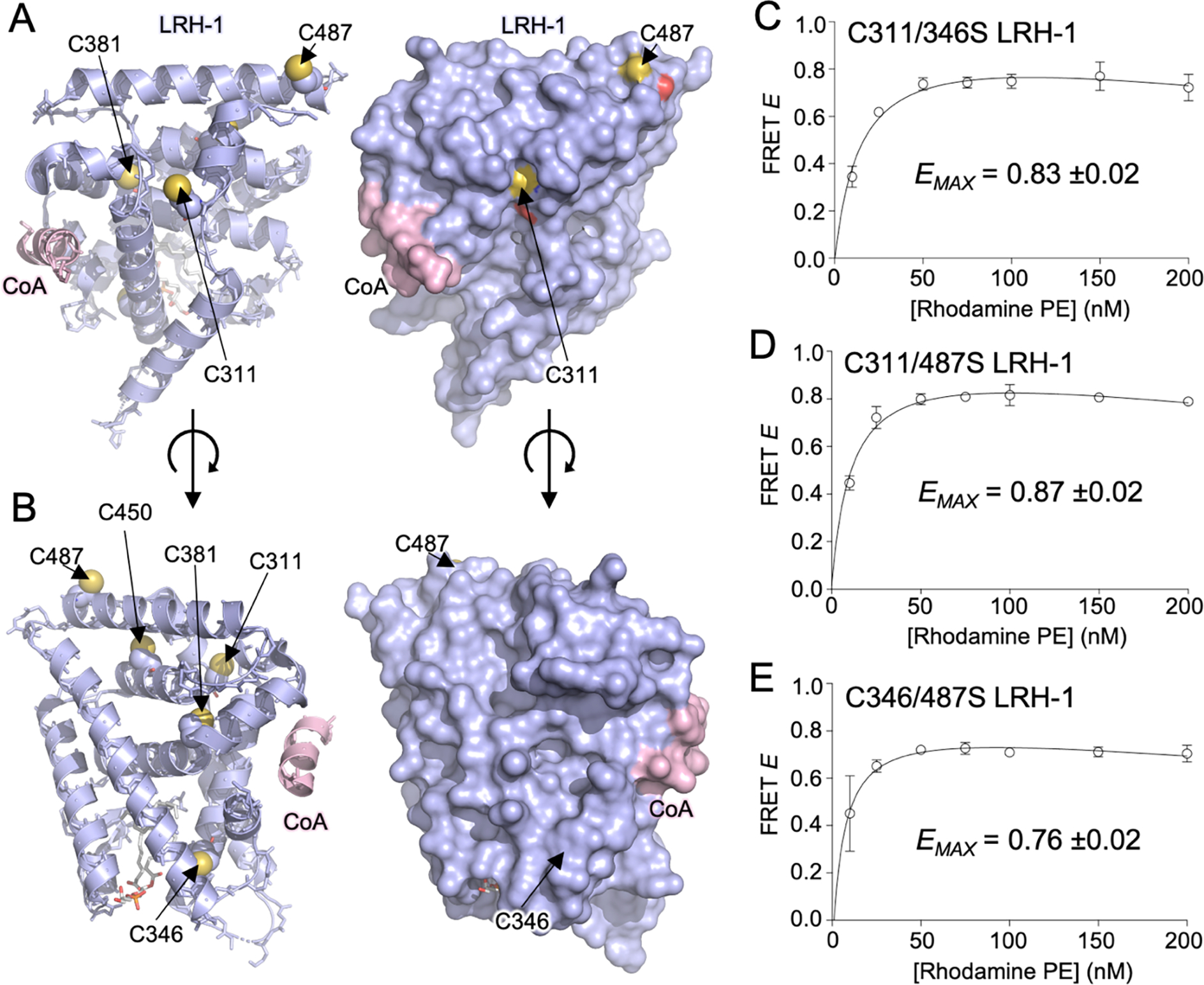
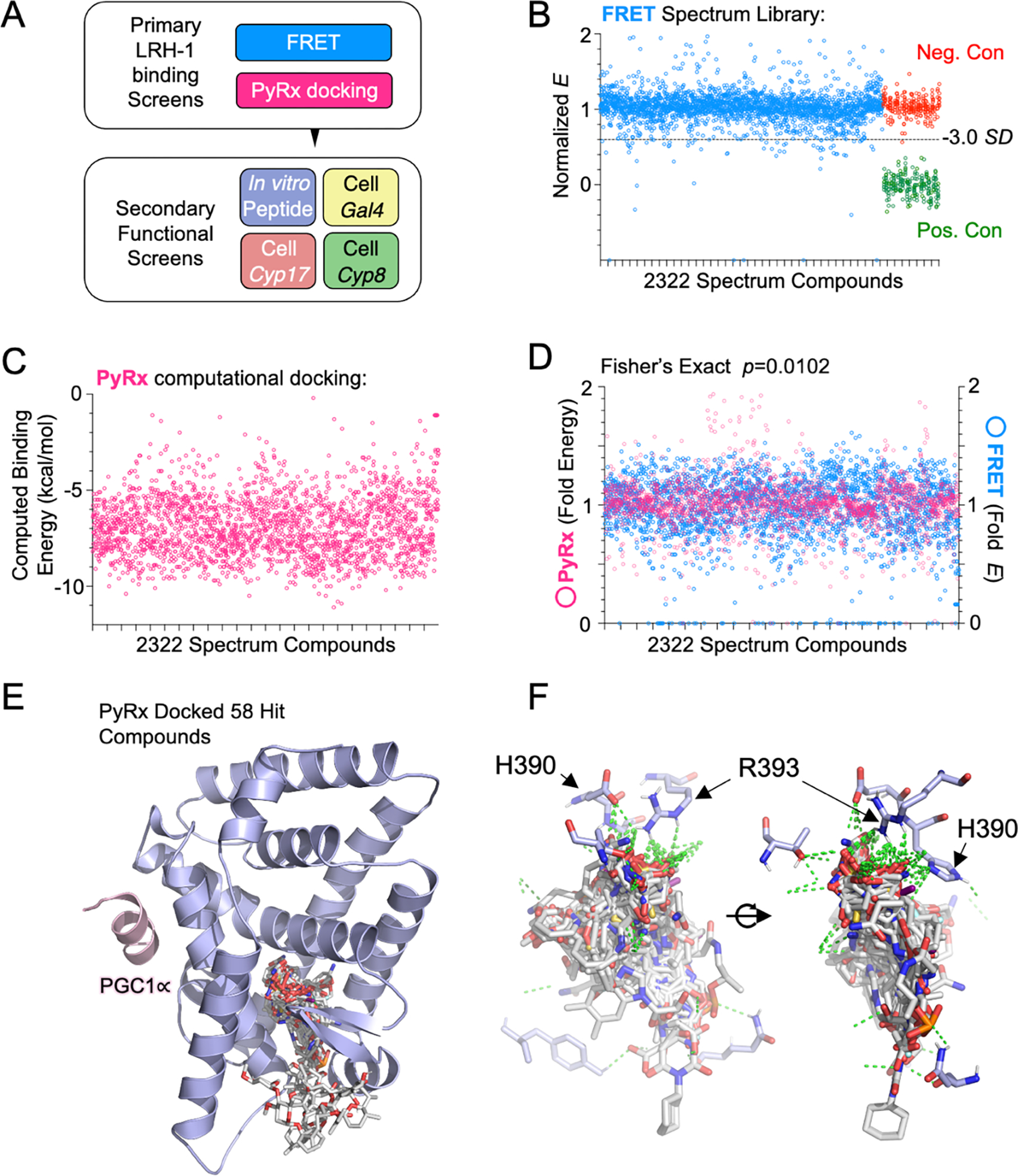
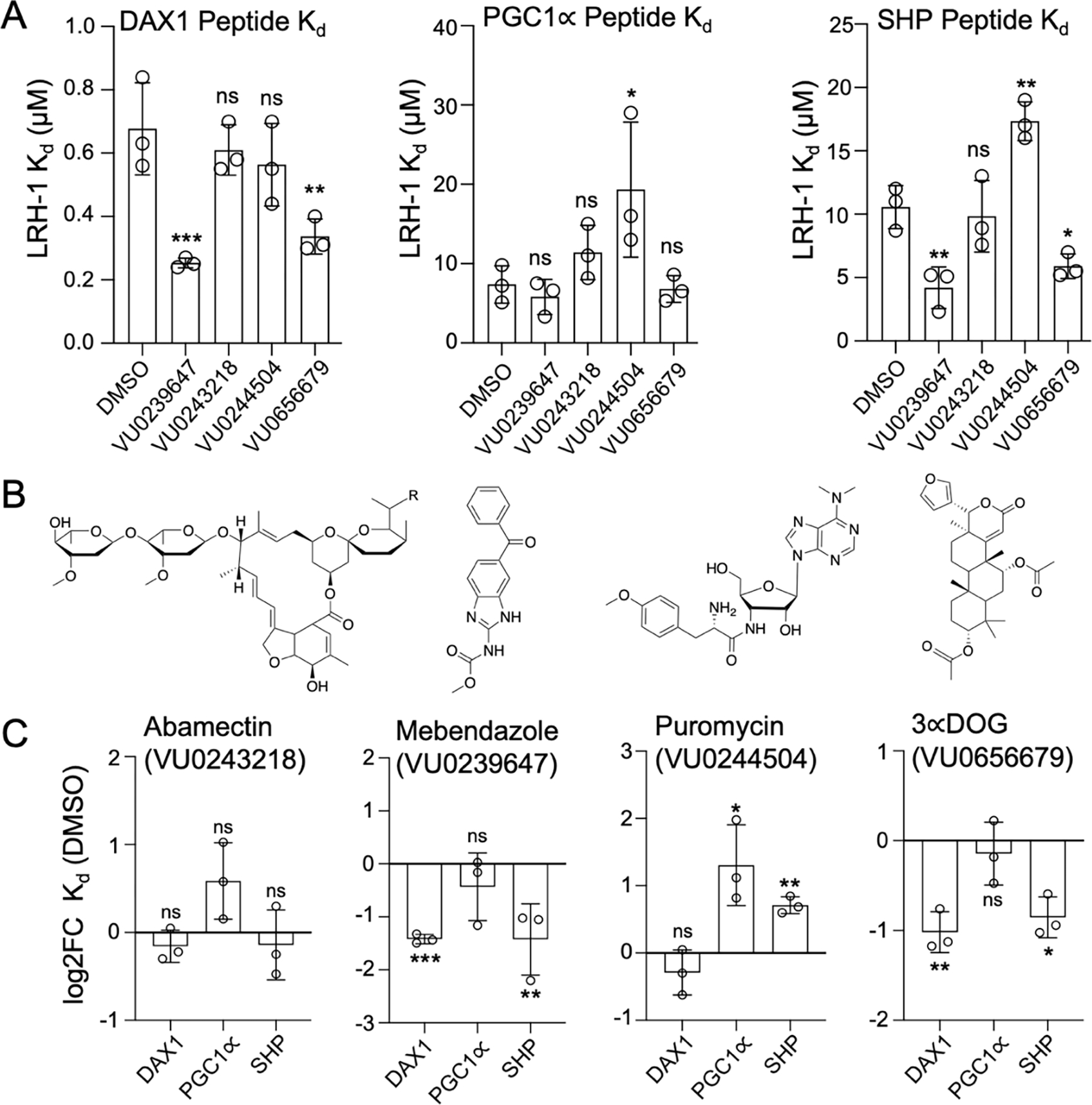
Nuclear receptor liver receptor homolog-1 (LRH-1, NR5A2) is a lipid-regulated transcription factor and an important drug target for several liver diseases. Advances toward LRH-1 therapeutics have been driven recently by structural biology, with fewer contributions from compound screening. Standard LRH-1 screens detect compound-induced interaction between LRH-1 and a transcriptional coregulator peptide, an approach that excludes compounds that regulate LRH-1 through alternative mechanisms. Here, we developed a FRET-based LRH-1 screen that simply detects compound binding to LRH-1, applying it to discover 58 new compounds that bind the canonical ligand-binding site in LRH-1 (2.5% hit rate), also supported by computational docking. Four independent functional screens identified 15 of these 58 compounds to also regulate LRH-1 function in vitro or in living cells. Although one of these 15 compounds, abamectin, directly binds LRH-1 and regulates full-length LRH-1 in cells, abamectin failed to regulate the isolated ligand-binding domain in standard coregulator peptide recruitment assays using PGC1α, DAX-1, or SHP. Abamectin treatment of human liver HepG2 cells selectively regulated endogenous LRH-1 ChIP-seq target genes and pathways associated with known LRH-1 functions in bile acid and cholesterol metabolism. Thus, the screen reported here can discover compounds not likely to have been identified in standard LRH-1 compound screens but which bind and regulate full-length LRH-1 in cells.
Figures


Similar articles
-
A novel heuristic of rigid docking scores positively correlates with full-length nuclear receptor LRH-1 regulation.Comput Struct Biotechnol J. 2024 Jul 30;23:3065-3080. doi: 10.1016/j.csbj.2024.07.021. eCollection 2024 Dec. Comput Struct Biotechnol J. 2024. PMID: 39185441 Free PMC article.
-
Bilirubin is a new ligand for nuclear receptor Liver Receptor Homolog-1.bioRxiv [Preprint]. 2024 Jun 1:2024.05.05.592606. doi: 10.1101/2024.05.05.592606. bioRxiv. 2024. PMID: 38853895 Free PMC article. Preprint.
-
Structure and Dynamics of the Liver Receptor Homolog 1-PGC1α Complex.Mol Pharmacol. 2017 Jul;92(1):1-11. doi: 10.1124/mol.117.108514. Epub 2017 Mar 31. Mol Pharmacol. 2017. PMID: 28363985 Free PMC article.
-
LRH-1: an orphan nuclear receptor involved in development, metabolism and steroidogenesis.Trends Cell Biol. 2004 May;14(5):250-60. doi: 10.1016/j.tcb.2004.03.008. Trends Cell Biol. 2004. PMID: 15130581 Review.
-
The Orphan Nuclear Receptors Steroidogenic Factor-1 and Liver Receptor Homolog-1: Structure, Regulation, and Essential Roles in Mammalian Reproduction.Physiol Rev. 2019 Apr 1;99(2):1249-1279. doi: 10.1152/physrev.00019.2018. Physiol Rev. 2019. PMID: 30810078 Review.
Cited by
-
Fatty Acid Mimetic Fragments as Liver Receptor Homologue-1 Modulators.ACS Pharmacol Transl Sci. 2025 Feb 12;8(3):673-678. doi: 10.1021/acsptsci.4c00734. eCollection 2025 Mar 14. ACS Pharmacol Transl Sci. 2025. PMID: 40109748
-
A novel heuristic of rigid docking scores positively correlates with full-length nuclear receptor LRH-1 regulation.Comput Struct Biotechnol J. 2024 Jul 30;23:3065-3080. doi: 10.1016/j.csbj.2024.07.021. eCollection 2024 Dec. Comput Struct Biotechnol J. 2024. PMID: 39185441 Free PMC article.
-
Fragment-Based Discovery of Drug-like LRH-1 Agonists.ACS Med Chem Lett. 2025 Mar 20;16(4):575-582. doi: 10.1021/acsmedchemlett.4c00604. eCollection 2025 Apr 10. ACS Med Chem Lett. 2025. PMID: 40236550
References
-
- Schoonjans K; Staels B; Auwerx J The Peroxisome Proliferator Activated Receptors (PPARs) and Their Effects on Lipid Metabolism and Adipocyte Differentiation. Biochimica et Biophysica Acta - Lipids and Lipid Metabolism. Elsevier B.V. July 26, 1996, pp 93–109. 10.1016/0005-2760(96)00066-5. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
