

Identification of D-arabinan-degrading enzymes in mycobacteria
- PMID: 37076525
- PMCID: PMC10115798
- DOI: 10.1038/s41467-023-37839-5
Identification of D-arabinan-degrading enzymes in mycobacteria
Abstract
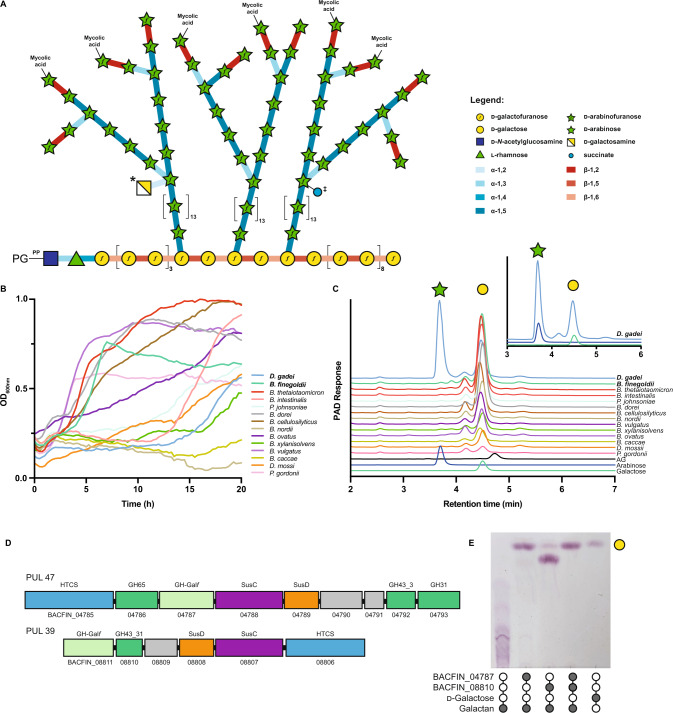
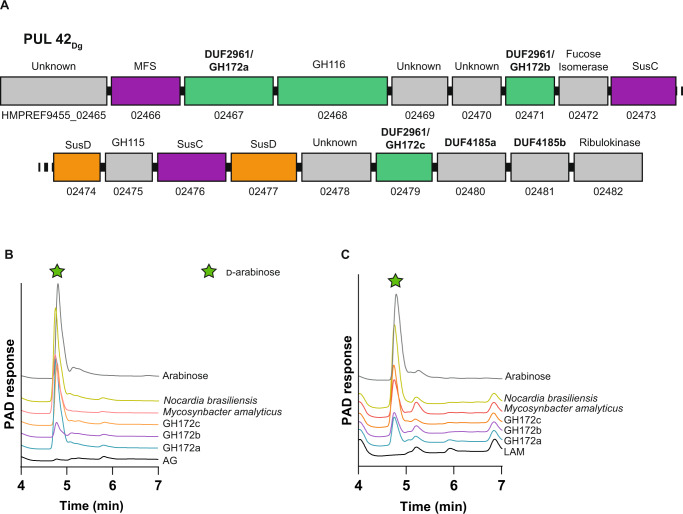
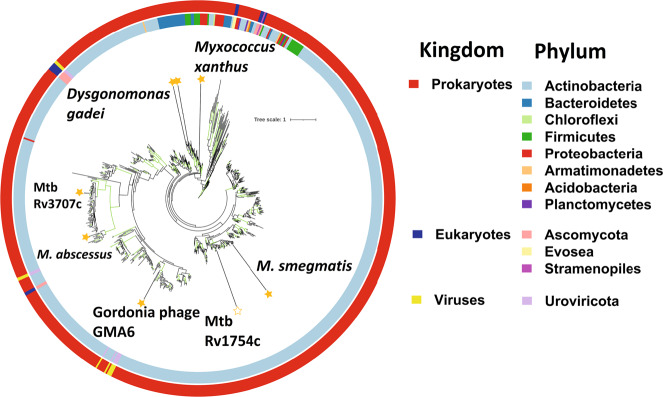
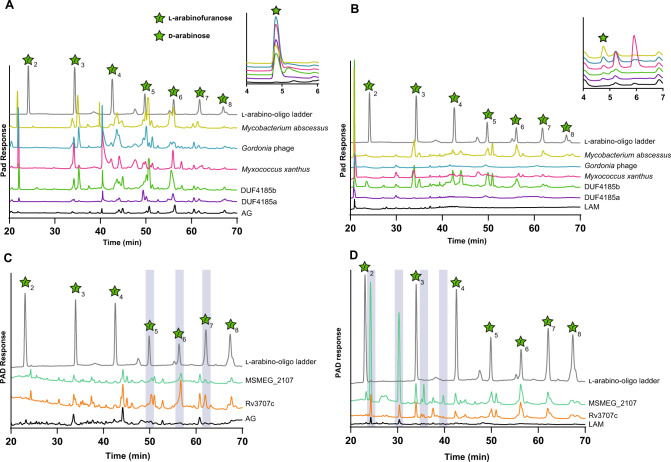
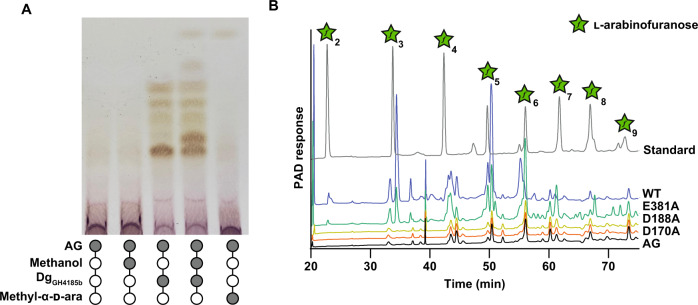
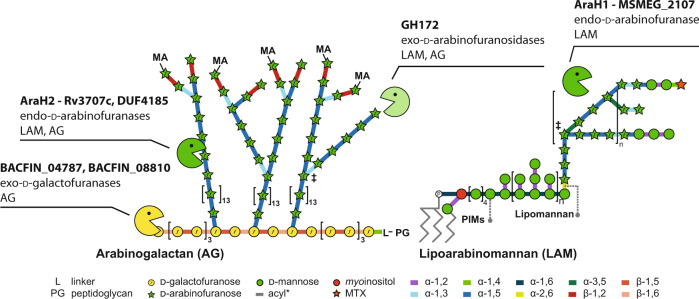
Bacterial cell growth and division require the coordinated action of enzymes that synthesize and degrade cell wall polymers. Here, we identify enzymes that cleave the D-arabinan core of arabinogalactan, an unusual component of the cell wall of Mycobacterium tuberculosis and other mycobacteria. We screened 14 human gut-derived Bacteroidetes for arabinogalactan-degrading activities and identified four families of glycoside hydrolases with activity against the D-arabinan or D-galactan components of arabinogalactan. Using one of these isolates with exo-D-galactofuranosidase activity, we generated enriched D-arabinan and used it to identify a strain of Dysgonomonas gadei as a D-arabinan degrader. This enabled the discovery of endo- and exo-acting enzymes that cleave D-arabinan, including members of the DUF2961 family (GH172) and a family of glycoside hydrolases (DUF4185/GH183) that display endo-D-arabinofuranase activity and are conserved in mycobacteria and other microbes. Mycobacterial genomes encode two conserved endo-D-arabinanases with different preferences for the D-arabinan-containing cell wall components arabinogalactan and lipoarabinomannan, suggesting they are important for cell wall modification and/or degradation. The discovery of these enzymes will support future studies into the structure and function of the mycobacterial cell wall.
© 2023. The Author(s).
Conflict of interest statement
Drs. Moynihan and Lowe are co-inventors on an unpublished patent application pertaining to some of the enzymes described in this manuscript. The remaining authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- BBSRCIAA-1544084 /BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/M011186/1 /BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- U01 GM125288/GM/NIGMS NIH HHS/United States
- R01 AI126592/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
