

Using quantitative methods to understand leaf epidermal development
- PMID: 37077990
- PMCID: PMC10097589
- DOI: 10.1017/qpb.2022.25
Using quantitative methods to understand leaf epidermal development
Abstract
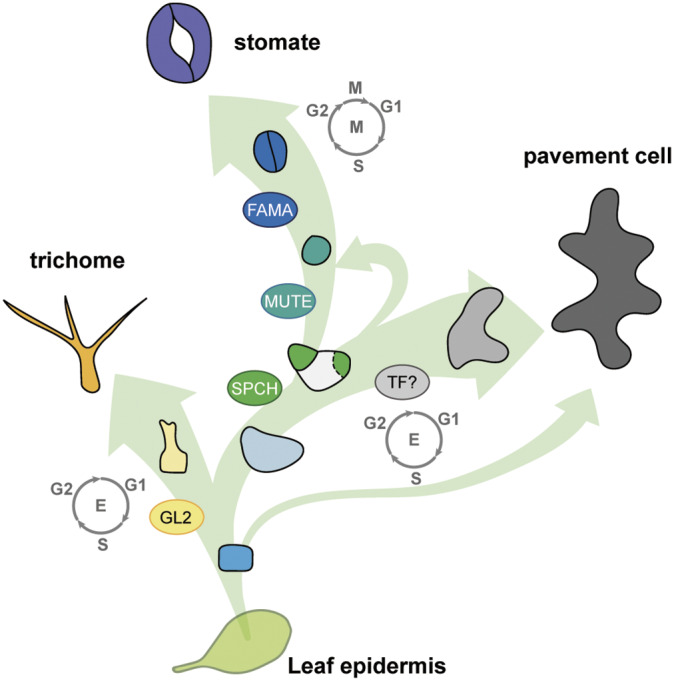
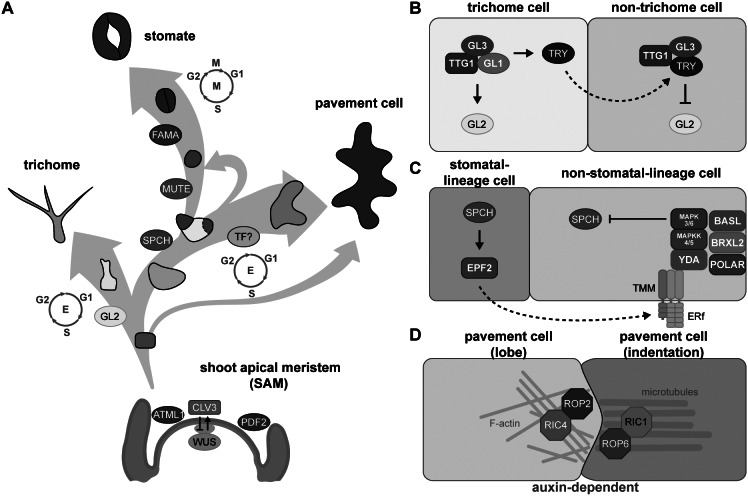
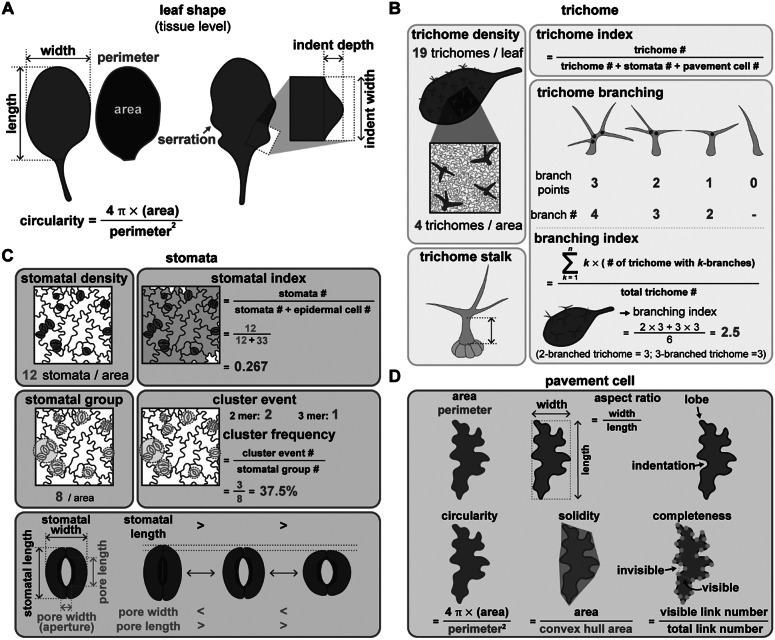
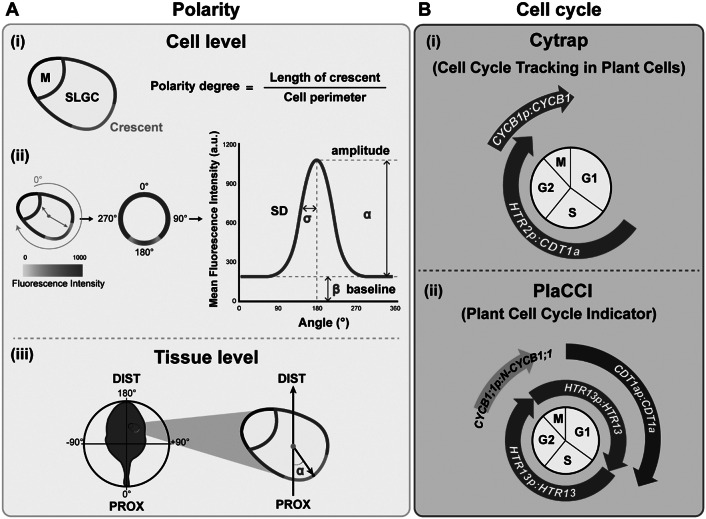
As the interface between plants and the environment, the leaf epidermis provides the first layer of protection against drought, ultraviolet light, and pathogen attack. This cell layer comprises highly coordinated and specialised cells such as stomata, pavement cells and trichomes. While much has been learned from the genetic dissection of stomatal, trichome and pavement cell formation, emerging methods in quantitative measurements that monitor cellular or tissue dynamics will allow us to further investigate cell state transitions and fate determination in leaf epidermal development. In this review, we introduce the formation of epidermal cell types in Arabidopsis and provide examples of quantitative tools to describe phenotypes in leaf research. We further focus on cellular factors involved in triggering cell fates and their quantitative measurements in mechanistic studies and biological patterning. A comprehensive understanding of how a functional leaf epidermis develops will advance the breeding of crops with improved stress tolerance.
Keywords: SLGC; cell state; leaf epidermal development; quantitative methods; stomata.
© The Author(s) 2022.
Conflict of interest statement
The authors declare none.
Figures





References
-
- Abe, M. , Katsumata, H. , Komeda, Y. , & Takahashi, T. (2003). Regulation of shoot epidermal cell differentiation by a pair of homeodomain proteins in Arabidopsis. Development (Cambridge, England), 130, 635–643. - PubMed
-
- Abe, T. , Thitamadee, S. , & Hashimoto, T. (2004). Microtubule defects and cell morphogenesis in the lefty1lefty2 tubulin mutant of Arabidopsis thaliana. Plant and Cell Physiology, 45, 211–220. - PubMed
-
- Altartouri, B. , Bidhendi, A. J. , Tani, T. , Suzuki, J. , Conrad, C. , Chebli, Y. , Liu, N. , Karunakaran, C. , Scarcelli, G. , & Geitmann, A. (2019). Pectin chemistry and cellulose crystallinity govern pavement cell morphogenesis in a multi-step mechanism. Plant Physiology, 181, 127–141. - PMC - PubMed
-
- Asaoka, M. , Ooe, M. , Gunji, S. , Milani, P. , Runel, G. , Horiguchi, G. , Hamant, O. , Sawa, S. , Tsukaya, H. , & Ferjani, A. (2021). Stem integrity in Arabidopsis thaliana requires a load-bearing epidermis. Development (Cambridge, England), 148, dev198028. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Research Materials