

APP deficiency and HTRA2 modulates PrPc proteostasis in human cancer cells
- PMID: 37082595
- PMCID: PMC10074928
- DOI: 10.1016/j.bbadva.2021.100035
APP deficiency and HTRA2 modulates PrPc proteostasis in human cancer cells
Abstract
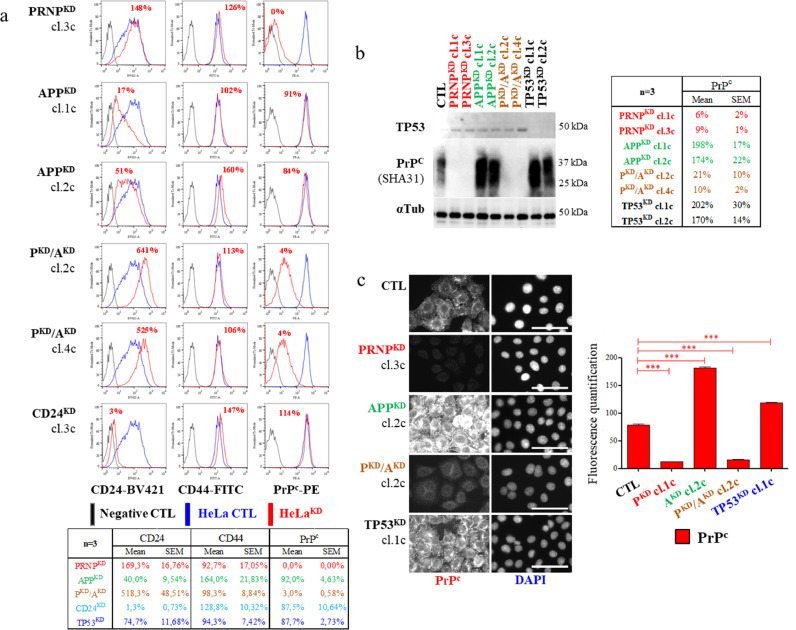
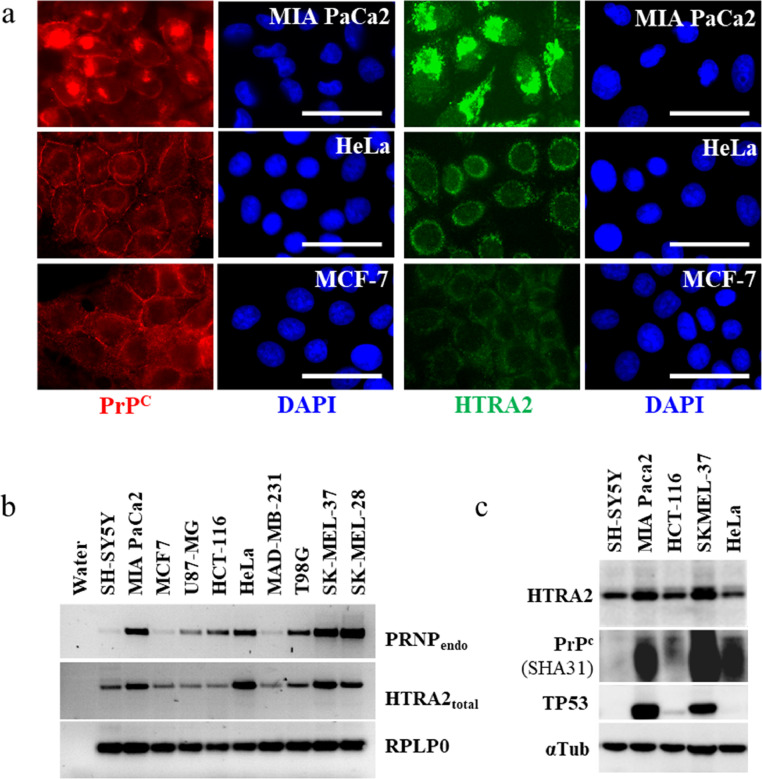
Cellular protein homeostasis (proteostasis) requires an accurate balance between protein biosynthesis, folding, and degradation, and its instability is causally related to human diseases and cancers. Here, we created numerous engineered cancer cell lines targeting APP (amyloid ß precursor protein) and/or PRNP (cellular prion) genes and we showed that APP knocking-down impaired PRNP mRNA level and vice versa, suggesting a link between their gene regulation. PRNPKD, APPKD and PRNPKD/APPKD HeLa cells encountered major difficulties to grow in a 3D tissue-like environment. Unexpectedly, we found a cytoplasmic accumulation of the PrPc protein without PRNP gene up regulation, in both APPKD and APPKO HeLa cells. Interestingly, APP and/or PRNP gene ablation enhanced the chaperone/serine protease HTRA2 gene expression, which is a protein processing quality factor involved in Alzheimer's disease. Importantly, HTRA2 gene silencing decreased PRNP mRNA level and lowered PrPc protein amounts, and conversely, HTRA2 overexpression increased PRNP gene regulation and enhanced membrane-anchored and cytoplasmic PrPc fractions. PrPc, APP and HTRA2 destabilized membrane-associated CD24 protein, suggesting changes in the lipid raft structure. Our data show for the first time that APP and the dual chaperone/serine protease HTRA2 protein could modulate PrPc proteostasis hampering cancer cell behavior.
Keywords: APP; CD24; CRISPR-Cas9; HTRA2; PRNP; PrPc; RNA interference; cancer cells; pEBVsiRNA; proteostasis.
© 2021 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
LinkOut - more resources
Full Text Sources
Research Materials
