

Diversification of ranunculaceous petals in shape supports a generalized model for plant lateral organ morphogenesis and evolution
- PMID: 37083529
- PMCID: PMC10121167
- DOI: 10.1126/sciadv.adf8049
Diversification of ranunculaceous petals in shape supports a generalized model for plant lateral organ morphogenesis and evolution
Abstract
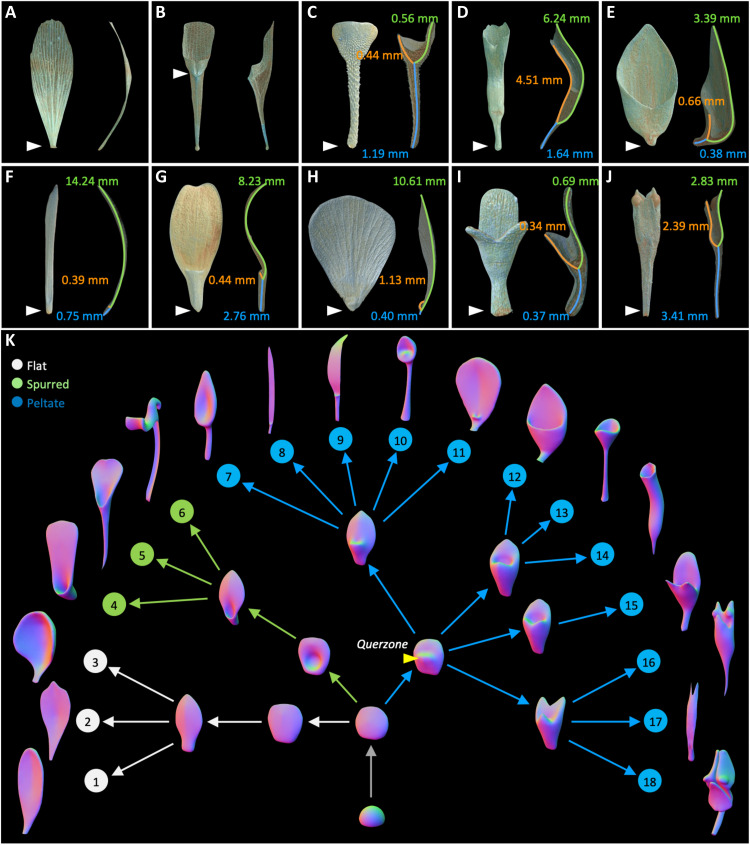
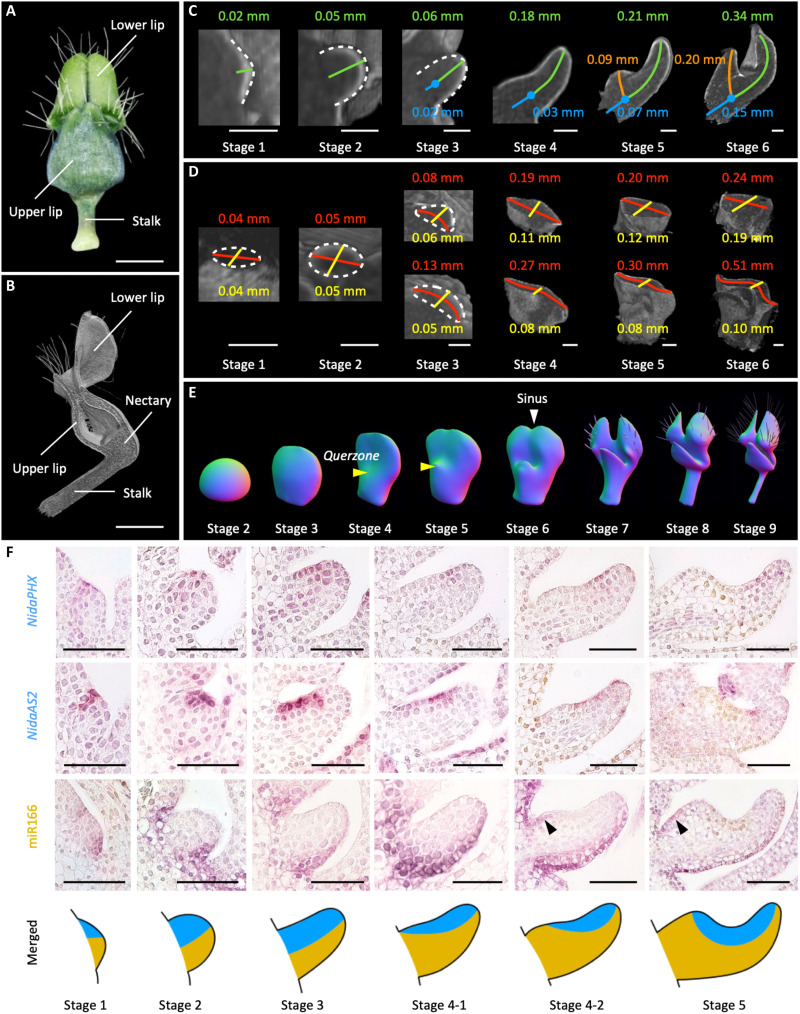
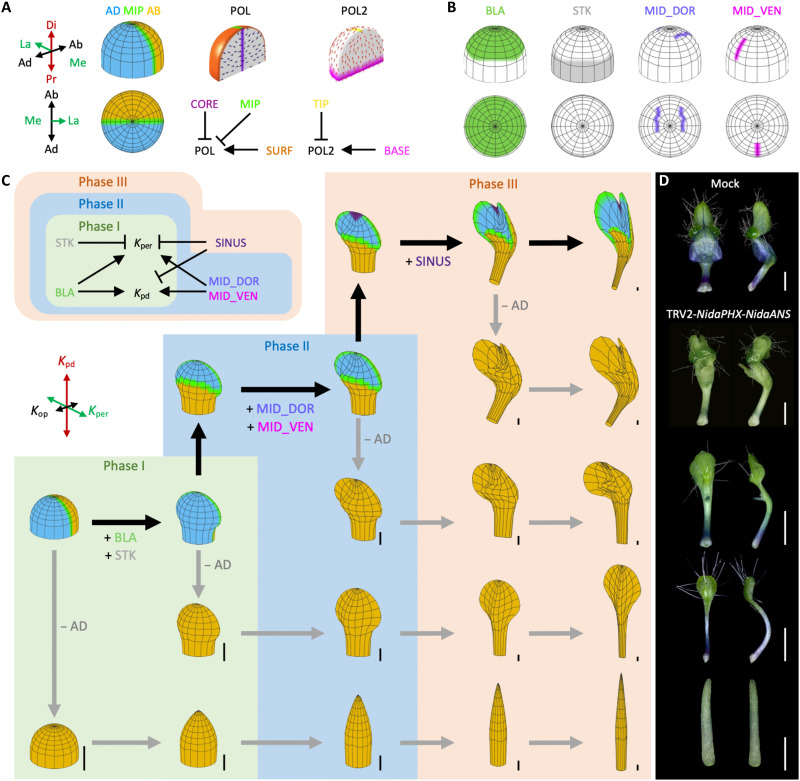
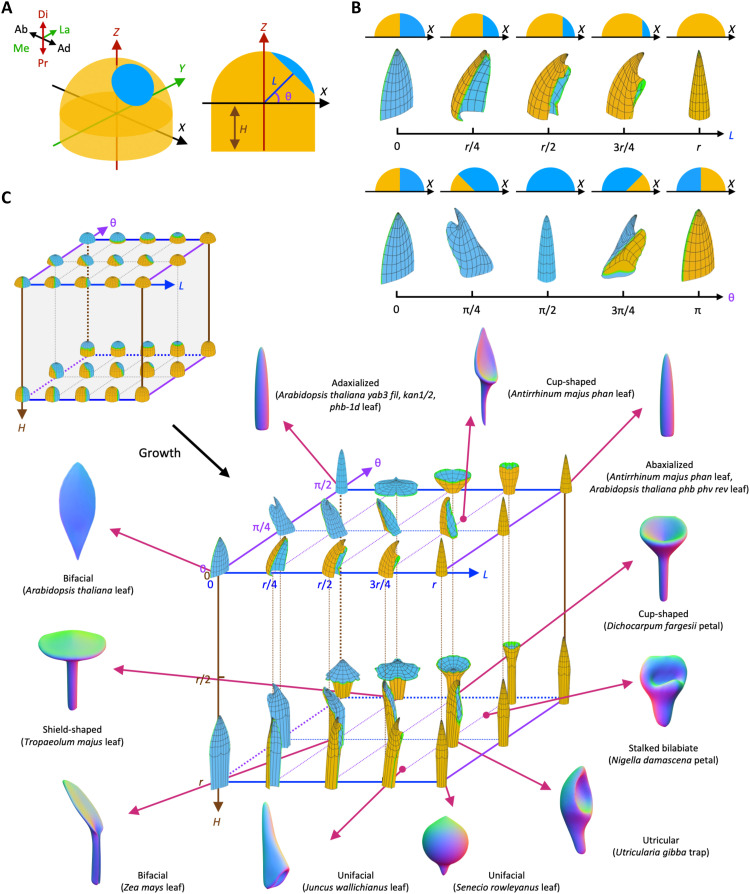
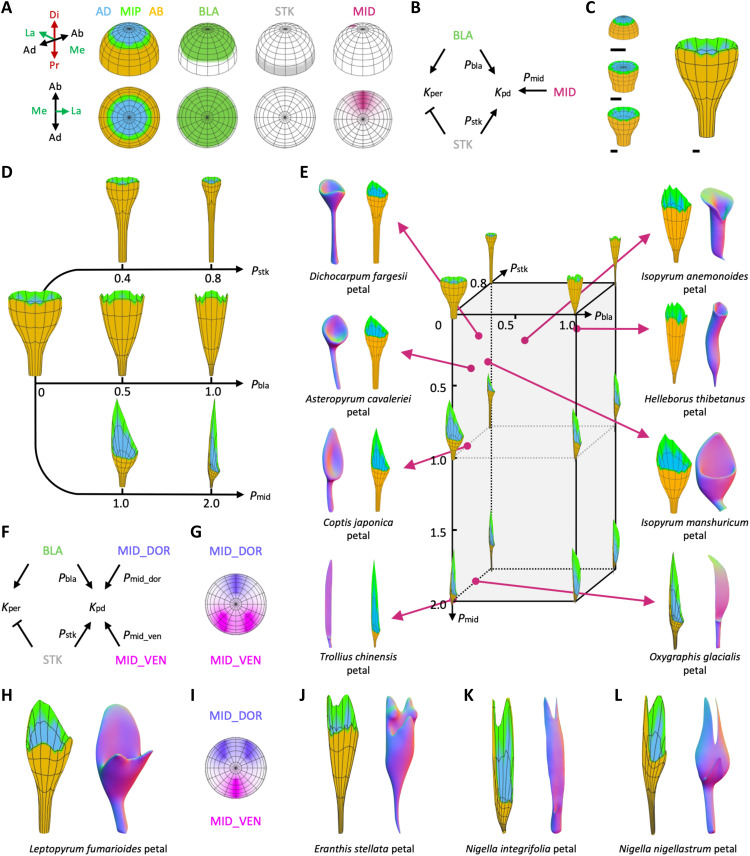
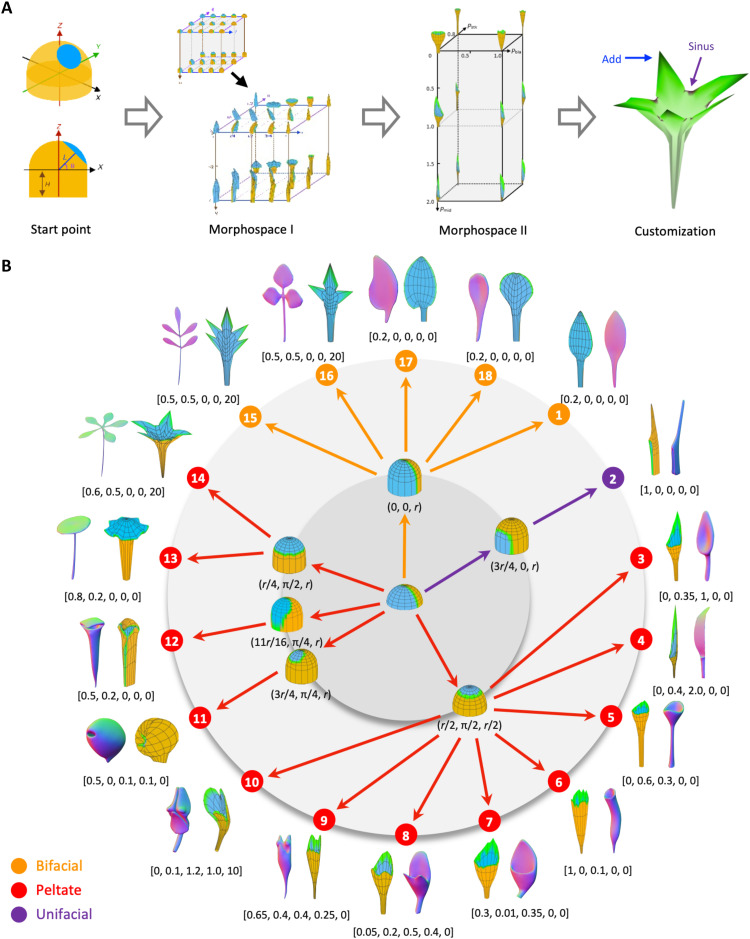
Peltate organs, such as the prey-capturing traps of carnivorous plants and nectary-bearing petals of ranunculaceous species, are widespread in nature and have intrigued and perplexed scientists for centuries. Shifts in the expression domains of adaxial/abaxial genes have been shown to control leaf peltation in some carnivorous plants, yet the mechanisms underlying the generation of other peltate organs remain unclear. Here, we show that formation of various peltate ranunculaceous petals was also caused by shifts in the expression domains of adaxial/abaxial genes, followed by differentiated regional growth sculpting the margins and/or other parts of the organs. By inducing parameters to specify the time, position, and degree of the shifts and growth, we further propose a generalized modeling system, through which various unifacial, bifacial, and peltate organs can be simulated. These results demonstrate the existence of a hierarchical morphospace system and pave the way to understand the mechanisms underlying plant organ diversification.
Figures
Similar articles
-
Adaxial-abaxial polarity: the developmental basis of leaf shape diversity.Genesis. 2014 Jan;52(1):1-18. doi: 10.1002/dvg.22728. Epub 2013 Dec 23. Genesis. 2014. PMID: 24281766 Review.
-
Developmental events leading to peltate leaf structure in Tropaeolum majus (Tropaeolaceae) are associated with expression domain changes of a YABBY gene.Dev Genes Evol. 2005 Jun;215(6):313-9. doi: 10.1007/s00427-005-0479-8. Epub 2005 Mar 25. Dev Genes Evol. 2005. PMID: 15791422
-
Leaf adaxial-abaxial polarity specification and lamina outgrowth: evolution and development.Plant Cell Physiol. 2012 Jul;53(7):1180-94. doi: 10.1093/pcp/pcs074. Epub 2012 May 21. Plant Cell Physiol. 2012. PMID: 22619472 Review.
-
Oriented cell division shapes carnivorous pitcher leaves of Sarracenia purpurea.Nat Commun. 2015 Mar 16;6:6450. doi: 10.1038/ncomms7450. Nat Commun. 2015. PMID: 25774486 Free PMC article.
-
Genetic framework for flattened leaf blade formation in unifacial leaves of Juncus prismatocarpus.Plant Cell. 2010 Jul;22(7):2141-55. doi: 10.1105/tpc.110.076927. Epub 2010 Jul 20. Plant Cell. 2010. PMID: 20647346 Free PMC article.
Cited by
-
A diffusible small-RNA-based Turing system dynamically coordinates organ polarity.Nat Plants. 2024 Mar;10(3):412-422. doi: 10.1038/s41477-024-01634-x. Epub 2024 Feb 26. Nat Plants. 2024. PMID: 38409292
-
Biregionally differentiated growth generates sharp apex and concave joints in leaves.Plant J. 2025 Jul;123(1):e70310. doi: 10.1111/tpj.70310. Plant J. 2025. PMID: 40616818 Free PMC article.
-
The GRAS transcription factor CsTL regulates tendril formation in cucumber.Plant Cell. 2024 Jul 31;36(8):2818-2833. doi: 10.1093/plcell/koae123. Plant Cell. 2024. PMID: 38630900 Free PMC article.
-
Evolution of petal patterning: blooming floral diversity at the microscale.New Phytol. 2025 Sep;247(6):2538-2556. doi: 10.1111/nph.70370. Epub 2025 Jul 8. New Phytol. 2025. PMID: 40629857 Free PMC article. Review.
-
Characterize the Complete Mitogenome of Semiaquilegia guangxiensis and Assess the Efficiency of the Mitochondrial Genes in Ranunculales Phylogeny.Ecol Evol. 2025 Mar 27;15(4):e71165. doi: 10.1002/ece3.71165. eCollection 2025 Apr. Ecol Evol. 2025. PMID: 40170818 Free PMC article.
References
-
- D. Franck, The morphological interpretation of epiascidiate leaves—An historical perspective. Bot. Rev. 42, 345–388 (1976).
-
- W. Troll, Morphologie der schieldförmigen blätter. Planta 17, 153–230 (1932).
-
- C. Thorogood, U. Bauer, S. Hiscock, Convergent and divergent evolution in carnivorous pitcher plant traps. New Phytol. 217, 1035–1041 (2018). - PubMed
-
- C. Erbar, S. Kusma, P. Leins, Development and interpretation of nectary organs in Ranunculaceae. Flora 194, 317–332 (1999).
-
- K. Kosuge, Petal evolution in Ranunculaceae. Plant Syst. Evol. 8, 185–191 (1994).
MeSH terms
LinkOut - more resources
Full Text Sources
