

Minor variations in multicellular life cycles have major effects on adaptation
- PMID: 37083675
- PMCID: PMC10156057
- DOI: 10.1371/journal.pcbi.1010698
Minor variations in multicellular life cycles have major effects on adaptation
Abstract
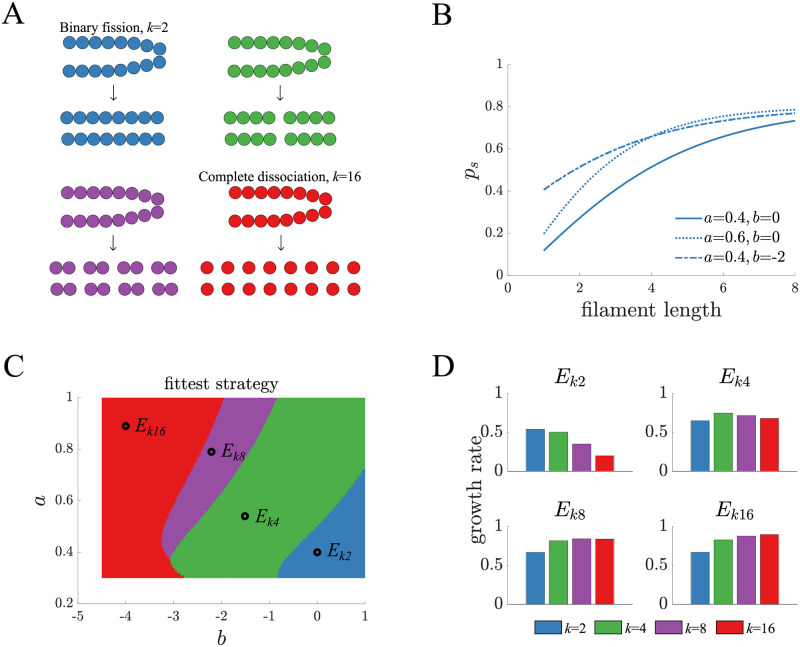
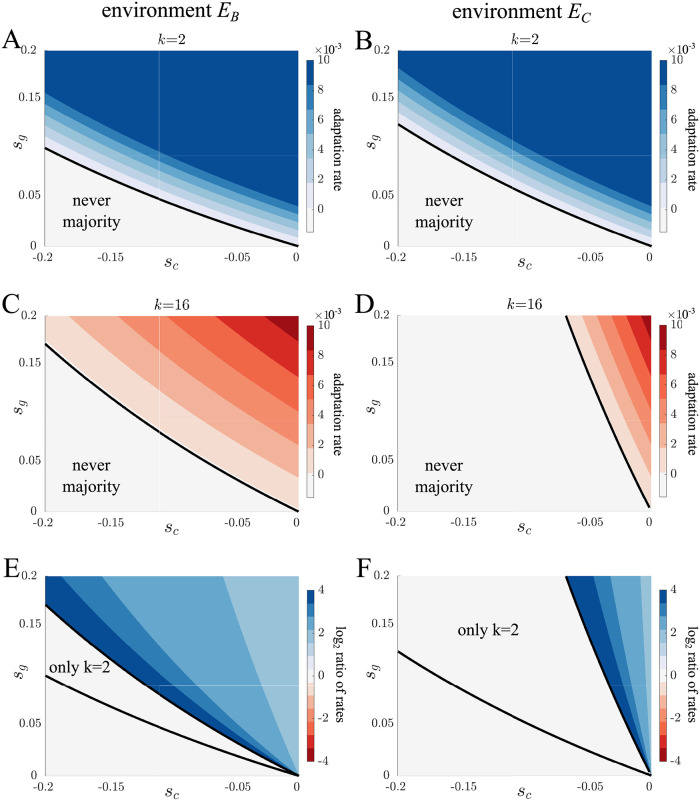
Multicellularity has evolved several independent times over the past hundreds of millions of years and given rise to a wide diversity of complex life. Recent studies have found that large differences in the fundamental structure of early multicellular life cycles can affect fitness and influence multicellular adaptation. Yet, there is an underlying assumption that at some scale or categorization multicellular life cycles are similar in terms of their adaptive potential. Here, we consider this possibility by exploring adaptation in a class of simple multicellular life cycles of filamentous organisms that only differ in one respect, how many daughter filaments are produced. We use mathematical models and evolutionary simulations to show that despite the similarities, qualitatively different mutations fix. In particular, we find that mutations with a tradeoff between cell growth and group survival, i.e. "selfish" or "altruistic" traits, spread differently. Specifically, altruistic mutations more readily spread in life cycles that produce few daughters while in life cycles producing many daughters either type of mutation can spread depending on the environment. Our results show that subtle changes in multicellular life cycles can fundamentally alter adaptation.
Copyright: © 2023 Isaksson et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
We the authors declare no competing interests.
Figures




References
-
- Schopf JW, Haugh BN, Molnar RE, Satterthwait DF. On the development of metaphytes and metazoans. Journal of Paleontology. 1973; p. 1–9.
-
- Smith JM, Szathmary E. The major transitions in evolution. Oxford University Press; 1997.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
