

Evolution of immunogenetic components encoding ultralong CDR H3
- PMID: 37084012
- PMCID: PMC10119515
- DOI: 10.1007/s00251-023-01305-9
Evolution of immunogenetic components encoding ultralong CDR H3
Abstract
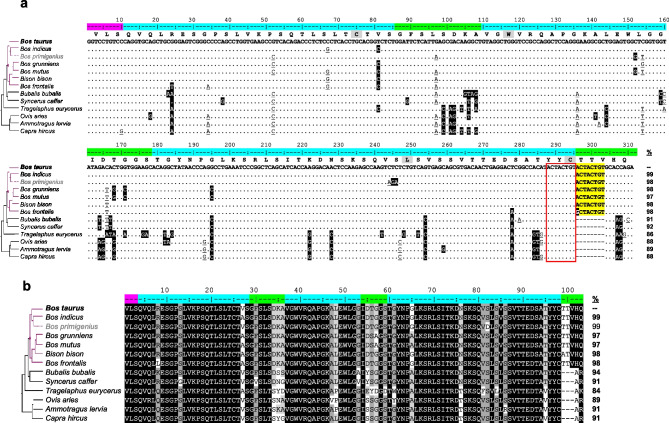
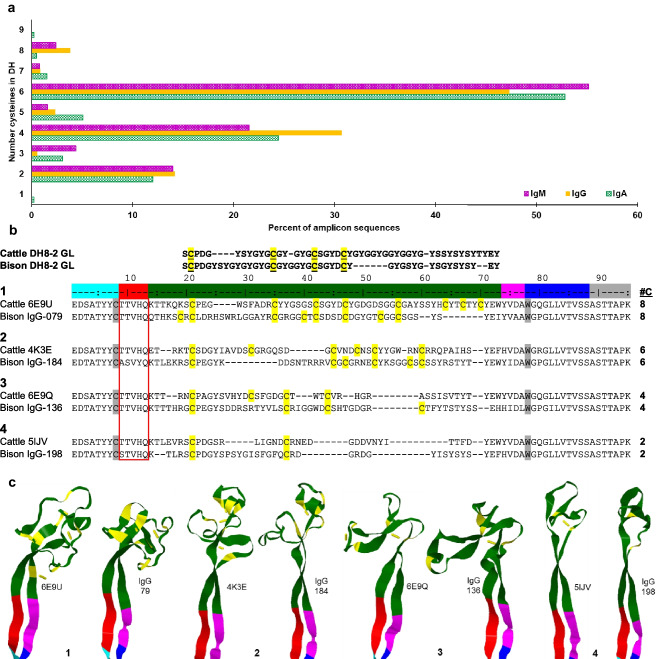
The genomes of most vertebrates contain many V, D, and J gene segments within their Ig loci to construct highly variable CDR3 sequences through combinatorial diversity. This nucleotide variability translates into an antibody population containing extensive paratope diversity. Cattle have relatively few functional VDJ gene segments, requiring innovative approaches for generating diversity like the use of ultralong-encoding IGHV and IGHD gene segments that yield dramatically elongated CDR H3. Unique knob and stalk microdomains create protracted paratopes, where the antigen-binding knob sits atop a long stalk, allowing the antibody to bind both surface and recessed antigen epitopes. We examined genomes of twelve species of Bovidae to determine when ultralong-encoding IGHV and IGHD gene segments evolved. We located the 8-bp duplication encoding the unique TTVHQ motif in ultralong IGHV segments in six Bovid species (cattle, zebu, wild yak, domestic yak, American bison, and domestic gayal), but we did not find evidence of the duplication in species beyond the Bos and Bison genera. Additionally, we analyzed mRNA from bison spleen and identified a rich repertoire of expressed ultralong CDR H3 antibody mRNA, suggesting that bison use ultralong IGHV transcripts in their host defense. We found ultralong-encoding IGHD gene segments in all the same species except domestic yak, but again not beyond the Bos and Bison clade. Thus, the duplication event leading to this ultralong-encoding IGHV gene segment and the emergence of the ultralong-encoding IGHD gene segment appears to have evolved in a common ancestor of the Bos and Bison genera 5-10 million years ago.
Keywords: Antibody; Cattle; Diversity segment; Evolution; IgH locus; Ultralong CDR H3.
© 2023. The Author(s), under exclusive licence to Springer-Verlag GmbH Germany, part of Springer Nature.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Antonacci R, Mineccia M, Lefranc MP, Ashmaoui HM, Lanave C, Piccinni B, Pesole G, Hassanane MS, Massari S, Ciccarese S. Expression and genomic analyses of Camelus dromedarius T cell receptor delta (TRD) genes reveal a variable domain repertoire enlargement due to CDR3 diversification and somatic mutation. Mol Immunol. 2011;48(12–13):1384–1396. doi: 10.1016/j.molimm.2011.03.011. - DOI - PubMed
-
- Burke MJ, Scott JNF, Minshull TC, Gao Z, Manfield I, Savic S, Stockley PG, Calabrese AN, Boyes J (2022) A bovine antibody possessing an ultralong complementarity-determining region CDRH3 targets a highly conserved epitope in sarbecovirus spike proteins. J Biol Chem 298(12):102624. 10.1016/j.jbc.2022.102624 - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
