

Death-associated protein kinase 3 regulates the myogenic reactivity of cerebral arteries
- PMID: 37084168
- PMCID: PMC10988501
- DOI: 10.1113/EP090631
Death-associated protein kinase 3 regulates the myogenic reactivity of cerebral arteries
Abstract
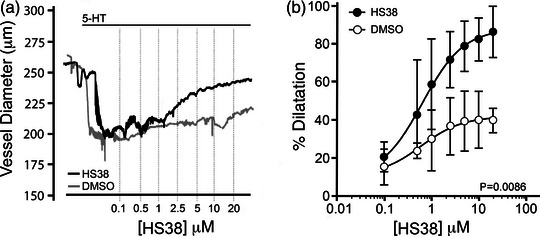
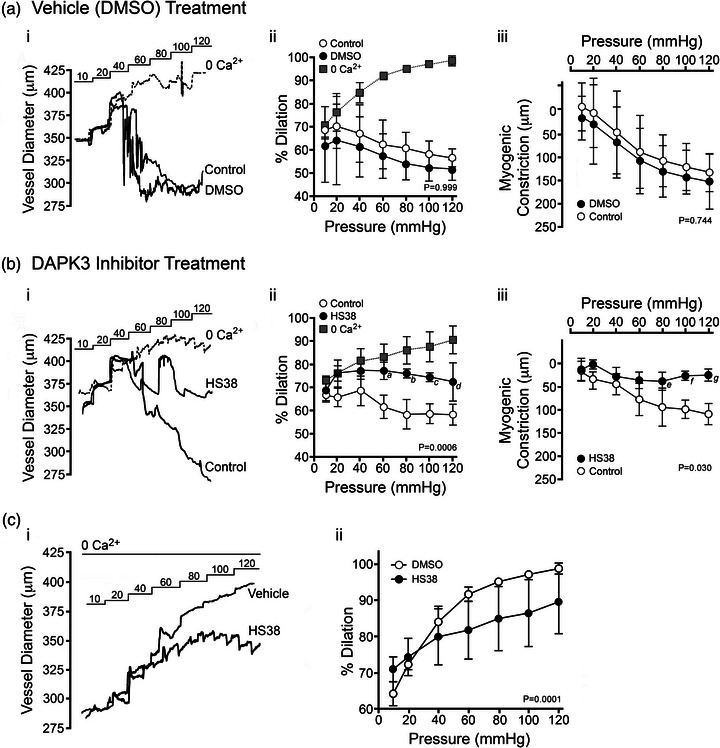
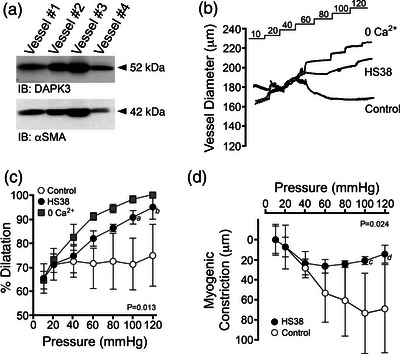
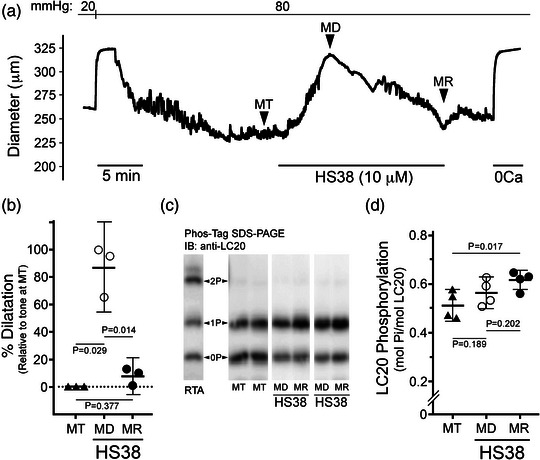
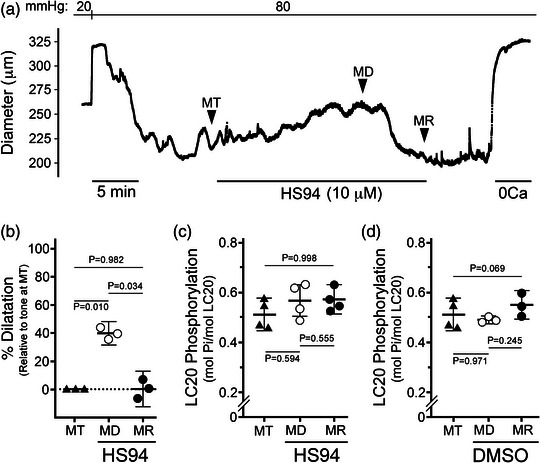
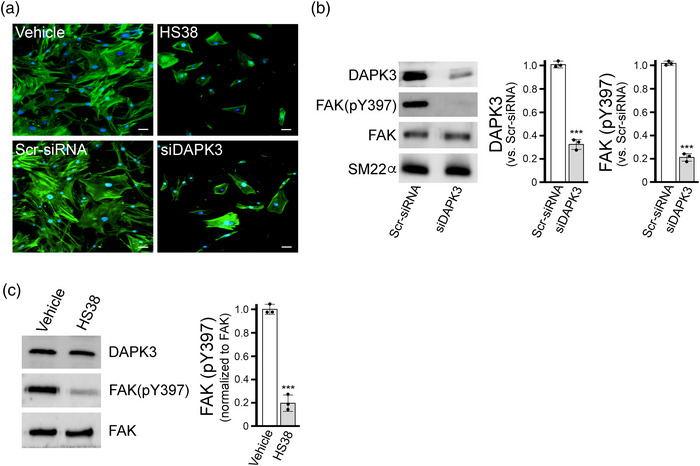
New findings: What is the central question of this study? DAPK3 contributes to the Ca2+ -sensitization of vascular smooth muscle contraction: does this protein kinase participate in the myogenic response of cerebral arteries? What is the main finding and its importance? Small molecule inhibitors of DAPK3 effectively block the myogenic responses of cerebral arteries. HS38-dependent changes to vessel constriction occur independent of LC20 phosphorylation, and therefore DAPK3 appears to operate via the actin cytoskeleton. A role for DAPK3 in the myogenic response was not previously reported, and the results support a potential new therapeutic target in the cerebrovascular system.
Abstract: The vascular smooth muscle (VSM) of resistance blood vessels is a target of intrinsic autoregulatory responses to increased intraluminal pressure, the myogenic response. In the brain, the myogenic reactivity of cerebral arteries is critical to homeostatic blood flow regulation. Here we provide the first evidence to link the death-associated protein kinase 3 (DAPK3) to the myogenic response of rat and human cerebral arteries. DAPK3 is a Ser/Thr kinase involved in Ca2+ -sensitization mechanisms of smooth muscle contraction. Ex vivo administration of a specific DAPK3 inhibitor (i.e., HS38) could attenuate vessel constrictions invoked by serotonin as well as intraluminal pressure elevation. The HS38-dependent dilatation was not associated with any change in myosin light chain (LC20) phosphorylation. The results suggest that DAPK3 does not regulate Ca2+ sensitization pathways during the myogenic response of cerebral vessels but rather operates to control the actin cytoskeleton. A slow return of myogenic tone was observed during the sustained ex vivo exposure of cerebral arteries to HS38. Recovery of tone was associated with greater LC20 phosphorylation that suggests intrinsic signalling compensation in response to attenuation of DAPK3 activity. Additional experiments with VSM cells revealed HS38- and siDAPK-dependent effects on the actin cytoskeleton and focal adhesion kinase phosphorylation status. The translational importance of DAPK3 to the human cerebral vasculature was noted, with robust expression of the protein kinase and significant HS38-dependent attenuation of myogenic reactivity found for human pial vessels.
Keywords: cerebrovascular; myogenic reactivity; vascular smooth muscle.
© 2023 The Authors. Experimental Physiology published by John Wiley & Sons Ltd on behalf of The Physiological Society.
Conflict of interest statement
J.A.M. is cofounder and has an equity position in Arch Biopartners Inc. T.A.J.H. is founder and has an equity position in Eydis Bio Inc. All other authors declare no conflicts of interest.
Figures
Similar articles
-
Ca2+ sensitization due to myosin light chain phosphatase inhibition and cytoskeletal reorganization in the myogenic response of skeletal muscle resistance arteries.J Physiol. 2013 Mar 1;591(5):1235-50. doi: 10.1113/jphysiol.2012.243576. Epub 2012 Dec 10. J Physiol. 2013. PMID: 23230233 Free PMC article.
-
α5-Integrin-mediated cellular signaling contributes to the myogenic response of cerebral resistance arteries.Biochem Pharmacol. 2015 Oct 1;97(3):281-91. doi: 10.1016/j.bcp.2015.08.088. Epub 2015 Aug 14. Biochem Pharmacol. 2015. PMID: 26278977
-
Abnormal myosin phosphatase targeting subunit 1 phosphorylation and actin polymerization contribute to impaired myogenic regulation of cerebral arterial diameter in the type 2 diabetic Goto-Kakizaki rat.J Cereb Blood Flow Metab. 2017 Jan;37(1):227-240. doi: 10.1177/0271678X15622463. Epub 2015 Dec 31. J Cereb Blood Flow Metab. 2017. PMID: 26721393 Free PMC article.
-
The role of actin filament dynamics in the myogenic response of cerebral resistance arteries.J Cereb Blood Flow Metab. 2013 Jan;33(1):1-12. doi: 10.1038/jcbfm.2012.144. Epub 2012 Oct 17. J Cereb Blood Flow Metab. 2013. PMID: 23072746 Free PMC article. Review.
-
Cellular signalling in arteriolar myogenic constriction: involvement of tyrosine phosphorylation pathways.Clin Exp Pharmacol Physiol. 2002 Jul;29(7):612-9. doi: 10.1046/j.1440-1681.2002.03698.x. Clin Exp Pharmacol Physiol. 2002. PMID: 12060106 Review.
Cited by
-
Organ-specific off-target effects of Pim/ZIP kinase inhibitors suggest lack of contractile Pim kinase activity in prostate, bladder, and vascular smooth muscle.Naunyn Schmiedebergs Arch Pharmacol. 2024 Feb;397(2):1219-1231. doi: 10.1007/s00210-023-02664-6. Epub 2023 Sep 1. Naunyn Schmiedebergs Arch Pharmacol. 2024. PMID: 37658212 Free PMC article.
References
-
- Abd‐Elrahman, K. S. , Colinas, O. , Walsh, E. J. , Zhu, H. L. , Campbell, C. M. , Walsh, M. P. , & Cole, W. C. (2017). Abnormal myosin phosphatase targeting subunit 1 phosphorylation and actin polymerization contribute to impaired myogenic regulation of cerebral arterial diameter in the type 2 diabetic Goto‐Kakizaki rat. Journal of Cerebral Blood Flow and Metabolism, 37(1), 227–240. - PMC - PubMed
-
- Borman, M. A. , MacDonald, J. A. , Muranyi, A. , Hartshorne, D. J. , & Haystead, T. A. (2002). Smooth muscle myosin phosphatase‐associated kinase induces Ca2+ sensitization via myosin phosphatase inhibition. Journal of Biological Chemistry, 277(26), 23441–23446. - PubMed
-
- Carlson, D. A. , Franke, A. S. , Weitzel, D. H. , Speer, B. L. , Hughes, P. F. , Hagerty, L. , Fortner, C. N. , Veal, J. M. , Barta, T. E. , Zieba, B. J. , Somlyo, A. V. , Sutherland, C. , Deng, J. T. , Walsh, M. P. , MacDonald, J. A. , & Haystead, T. A. (2013). Fluorescence linked enzyme chemoproteomic strategy for discovery of a potent and selective DAPK1 and ZIPK inhibitor. ACS Chemical Biology, 8(12), 2715–2723. - PMC - PubMed
-
- Carlson, D. A. , Singer, M. R. , Sutherland, C. , Redondo, C. , Alexander, L. T. , Hughes, P. F. , Knapp, S. , Gurley, S. B. , Sparks, M. A. , MacDonald, J. A. , & Haystead, T. A. J. (2018). Targeting Pim Kinases and DAPK3 to control hypertension. Cell Chemical Biology, 25(10), 1195–1207.e32. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous