

UDP-glucosyltransferase OsUGT75A promotes submergence tolerance during rice seed germination
- PMID: 37085517
- PMCID: PMC10121563
- DOI: 10.1038/s41467-023-38085-5
UDP-glucosyltransferase OsUGT75A promotes submergence tolerance during rice seed germination
Abstract
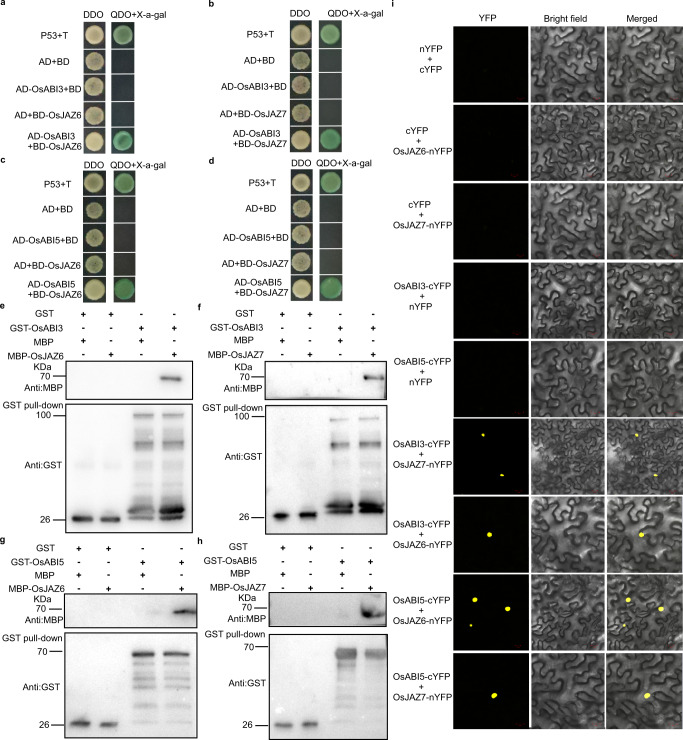
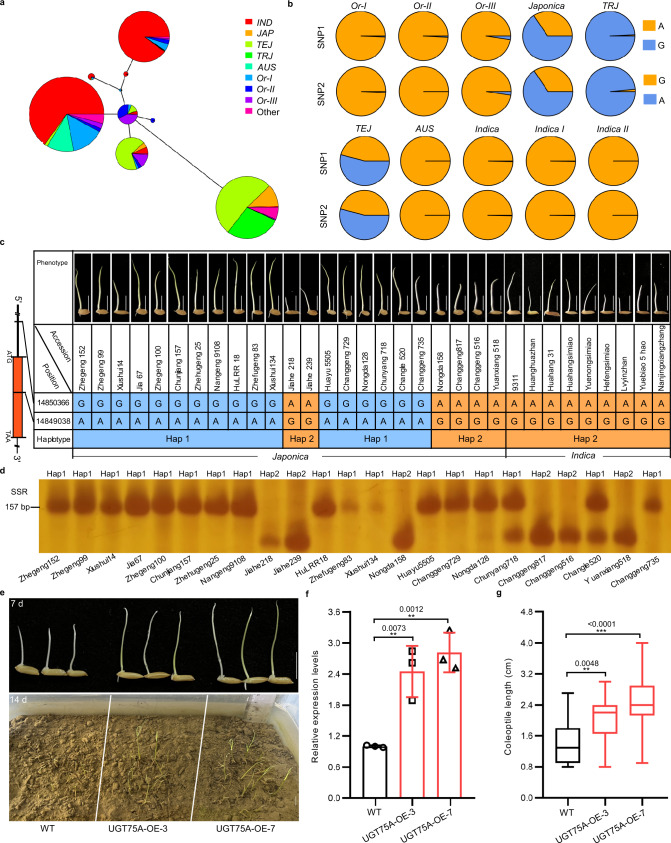
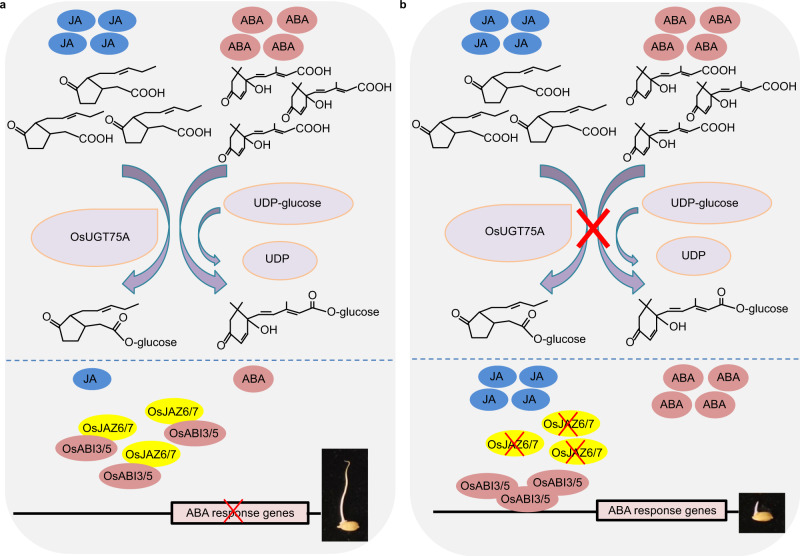
Submergence stress represents a major obstacle limiting the application of direct seeding in rice cultivation. Under flooding conditions, coleoptile elongation can function as an escape strategy that contributes to submergence tolerance during seed germination in rice; however, the underlying molecular bases have yet to be fully determined. Herein, we report that natural variation of rice coleoptile length subjected to submergence is determined by the glucosyltransferase encoding gene OsUGT75A. OsUGT75A regulates coleoptile length via decreasing free abscisic acid (ABA) and jasmonic acid (JA) levels by promoting glycosylation of these two phytohormones under submergence. Moreover, we find that OsUGT75A accelerates coleoptile length through mediating the interactions between JASMONATE ZIMDOMAIN (OsJAZ) and ABSCISIC ACID-INSENSITIVE (OsABI) proteins. Last, we reveal the origin of the haplotype that contributes to coleoptile length in response to submergence and transferring this haplotype to indica rice can enhance coleoptile length in submergence conditions. Thus, we propose that OsUGT75A is a useful target in breeding of rice varieties suitable for direct seeding cultivation.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





Similar articles
-
OsNAC3 regulates seed germination involving abscisic acid pathway and cell elongation in rice.New Phytol. 2024 Jan;241(2):650-664. doi: 10.1111/nph.19362. Epub 2023 Oct 31. New Phytol. 2024. PMID: 37908121
-
Promoter Architecture and Transcriptional Regulation of Genes Upregulated in Germination and Coleoptile Elongation of Diverse Rice Genotypes Tolerant to Submergence.Front Genet. 2021 Mar 16;12:639654. doi: 10.3389/fgene.2021.639654. eCollection 2021. Front Genet. 2021. PMID: 33796132 Free PMC article.
-
bZIP72 promotes submerged rice seed germination and coleoptile elongation by activating ADH1.Plant Physiol Biochem. 2021 Dec;169:112-118. doi: 10.1016/j.plaphy.2021.11.005. Epub 2021 Nov 6. Plant Physiol Biochem. 2021. PMID: 34775177
-
How does rice cope with too little oxygen during its early life?New Phytol. 2021 Jan;229(1):36-41. doi: 10.1111/nph.16395. Epub 2020 Jan 28. New Phytol. 2021. PMID: 31880324 Review.
-
The Molecular Regulatory Pathways and Metabolic Adaptation in the Seed Germination and Early Seedling Growth of Rice in Response to Low O2 Stress.Plants (Basel). 2020 Oct 14;9(10):1363. doi: 10.3390/plants9101363. Plants (Basel). 2020. PMID: 33066550 Free PMC article. Review.
Cited by
-
Advances in seed hypoxia research.Plant Physiol. 2024 Dec 23;197(1):kiae556. doi: 10.1093/plphys/kiae556. Plant Physiol. 2024. PMID: 39471319 Free PMC article.
-
Upregulation of an IAA-Glucosyltransferase OsIAGLU in Rice (Oryza sativa L.) Impairs Root Gravitropism by Disrupting Starch Granule Homeostasis.Plants (Basel). 2025 May 21;14(10):1557. doi: 10.3390/plants14101557. Plants (Basel). 2025. PMID: 40431120 Free PMC article.
-
Identification of Genomic Regions for Deep-Water Resistance in Rice for Efficient Weed Control with Reduced Herbicide Use.Rice (N Y). 2023 Nov 25;16(1):53. doi: 10.1186/s12284-023-00671-y. Rice (N Y). 2023. PMID: 38006407 Free PMC article.
-
Apple Glycosyltransferase MdUGT73AR4 Glycosylates ABA to Regulate Stomatal Movement Involved in Drought Stress.Int J Mol Sci. 2024 May 23;25(11):5672. doi: 10.3390/ijms25115672. Int J Mol Sci. 2024. PMID: 38891859 Free PMC article.
-
Differential adaptations of japonica rice to submergence stress during the tillering stage under various seedling cultivation and transplanting methods.Front Plant Sci. 2025 Jun 19;16:1607055. doi: 10.3389/fpls.2025.1607055. eCollection 2025. Front Plant Sci. 2025. PMID: 40612614 Free PMC article.
References
-
- Sasaki T. Studies on breeding for the germinability at low temperature of rice varieties adapted to direct sowing cultivation in flooded paddy field in cool region. Rep. Hokkaido Pref. Agric. Exp. Sta. 1974;24:1–90.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
