

Extracellular matrix assembly stress initiates Drosophila central nervous system morphogenesis
- PMID: 37086718
- PMCID: PMC10390342
- DOI: 10.1016/j.devcel.2023.03.019
Extracellular matrix assembly stress initiates Drosophila central nervous system morphogenesis
Abstract
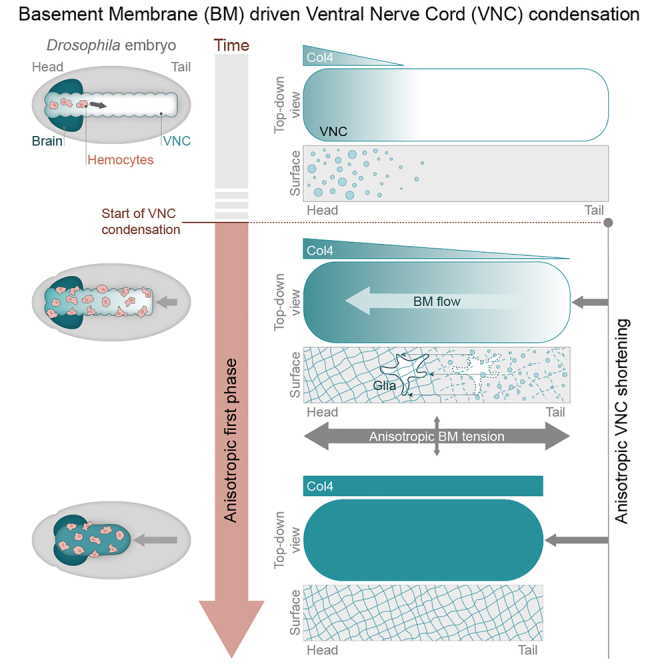
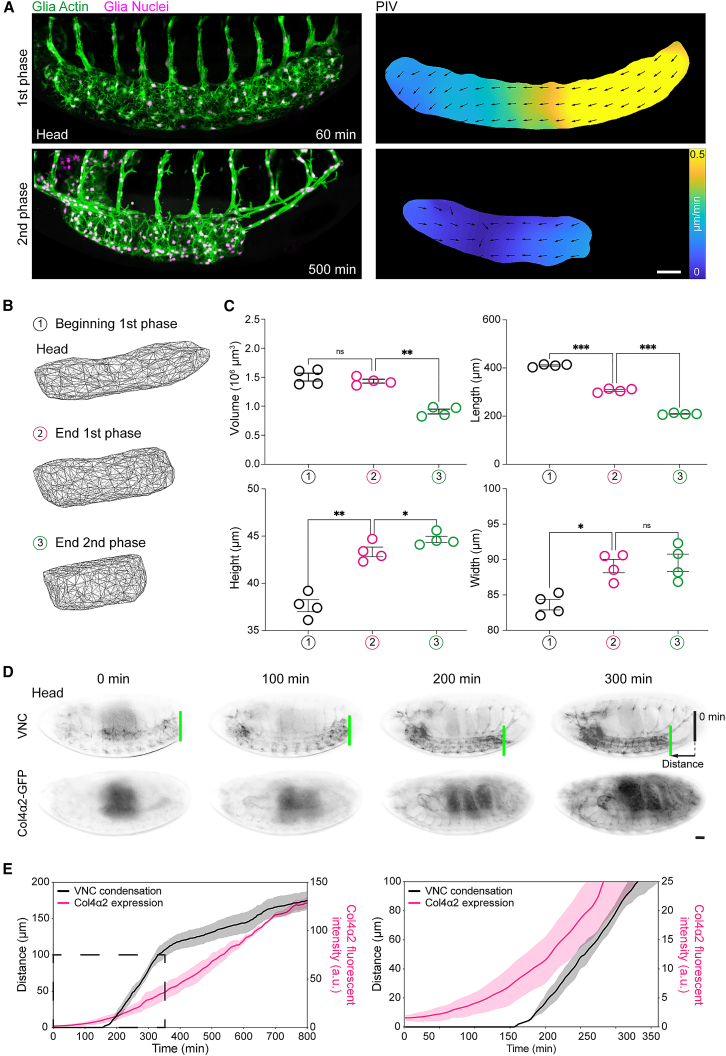
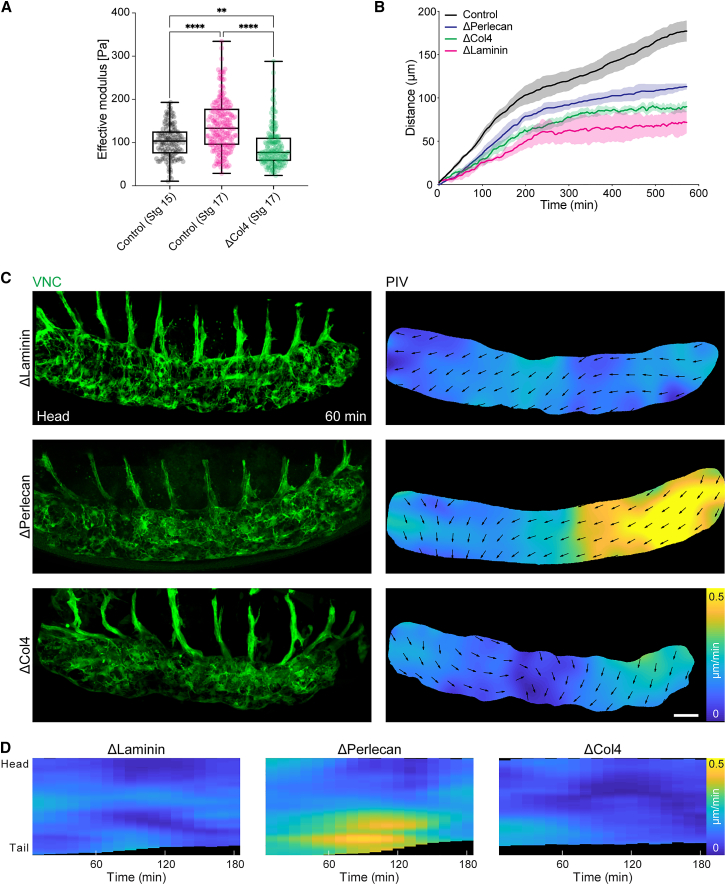
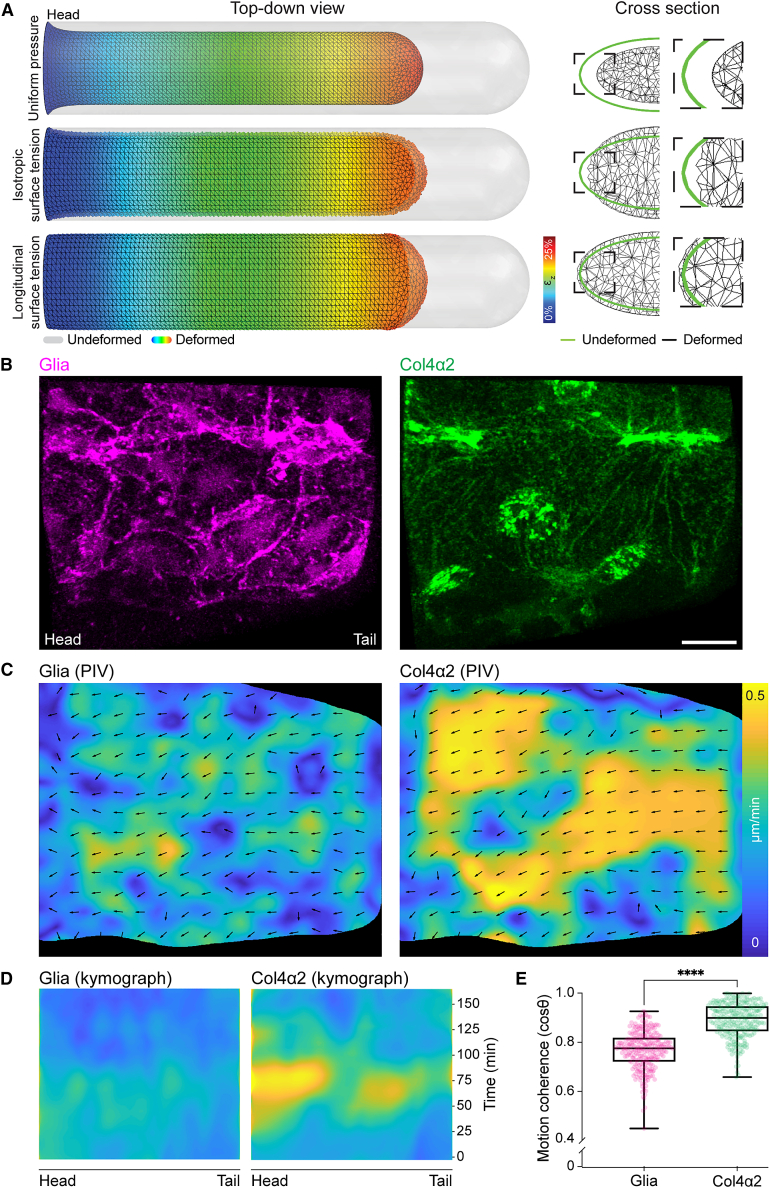
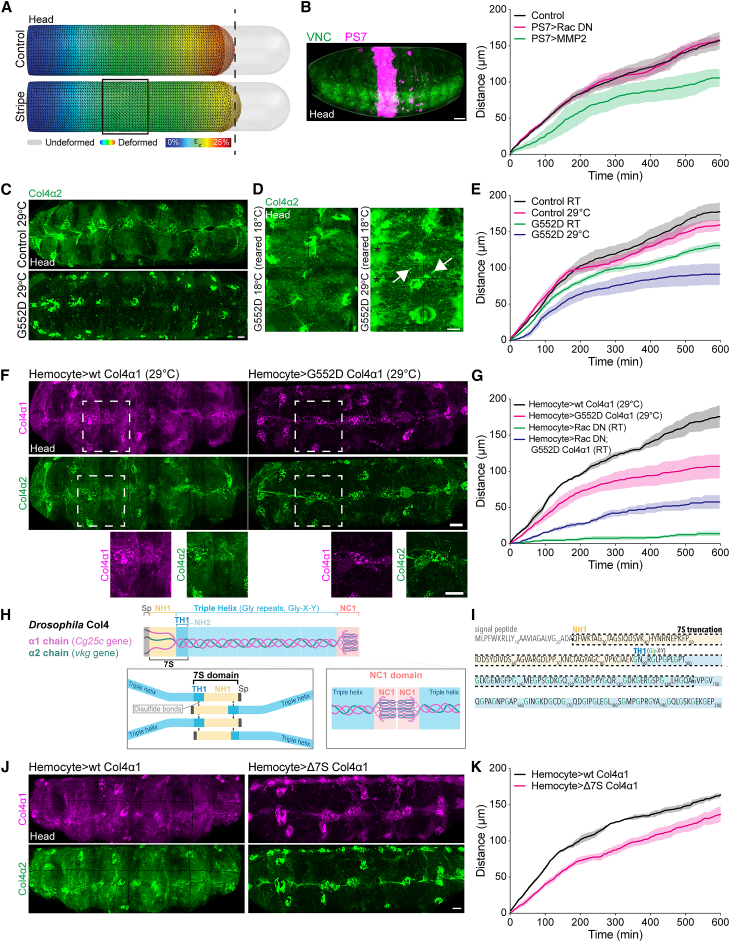
Forces controlling tissue morphogenesis are attributed to cellular-driven activities, and any role for extracellular matrix (ECM) is assumed to be passive. However, all polymer networks, including ECM, can develop autonomous stresses during their assembly. Here, we examine the morphogenetic function of an ECM before reaching homeostatic equilibrium by analyzing de novo ECM assembly during Drosophila ventral nerve cord (VNC) condensation. Asymmetric VNC shortening and a rapid decrease in surface area correlate with the exponential assembly of collagen IV (Col4) surrounding the tissue. Concomitantly, a transient developmentally induced Col4 gradient leads to coherent long-range flow of ECM, which equilibrates the Col4 network. Finite element analysis and perturbation of Col4 network formation through the generation of dominant Col4 mutations that affect assembly reveal that VNC morphodynamics is partially driven by a sudden increase in ECM-driven surface tension. These data suggest that ECM assembly stress and associated network instabilities can actively participate in tissue morphogenesis.
Keywords: Drosophila; basement membrane; central nervous system; collagen IV; embryonic development; extracellular matix; morphogenesis; surface tension.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Getting reshaped by the building of your substrate: Extracellular matrix assembly joins the morphogenesis toolkit.Dev Cell. 2023 May 22;58(10):823-824. doi: 10.1016/j.devcel.2023.04.017. Dev Cell. 2023. PMID: 37220739
References
-
- Gilbert S.F. Sixth Edition. Sinauer Associates; 2000. Developmental Biology.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
