

Long term worsening of amyloid pathology, cerebral function, and cognition after a single inoculation of beta-amyloid seeds with Osaka mutation
- PMID: 37087498
- PMCID: PMC10122826
- DOI: 10.1186/s40478-023-01559-0
Long term worsening of amyloid pathology, cerebral function, and cognition after a single inoculation of beta-amyloid seeds with Osaka mutation
Abstract
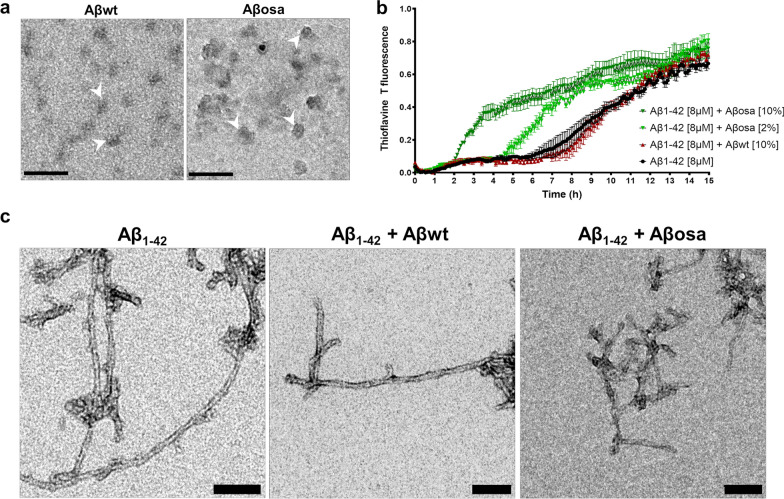
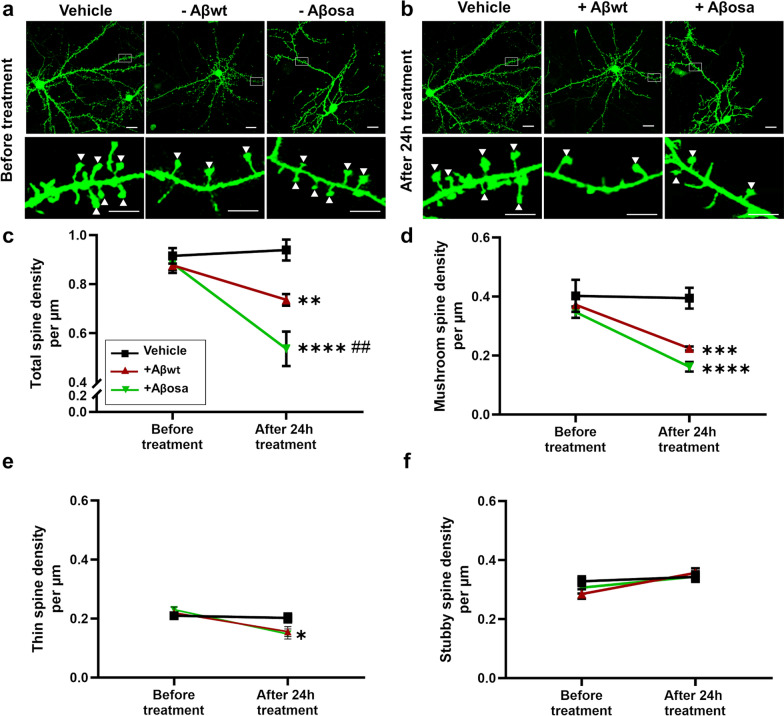
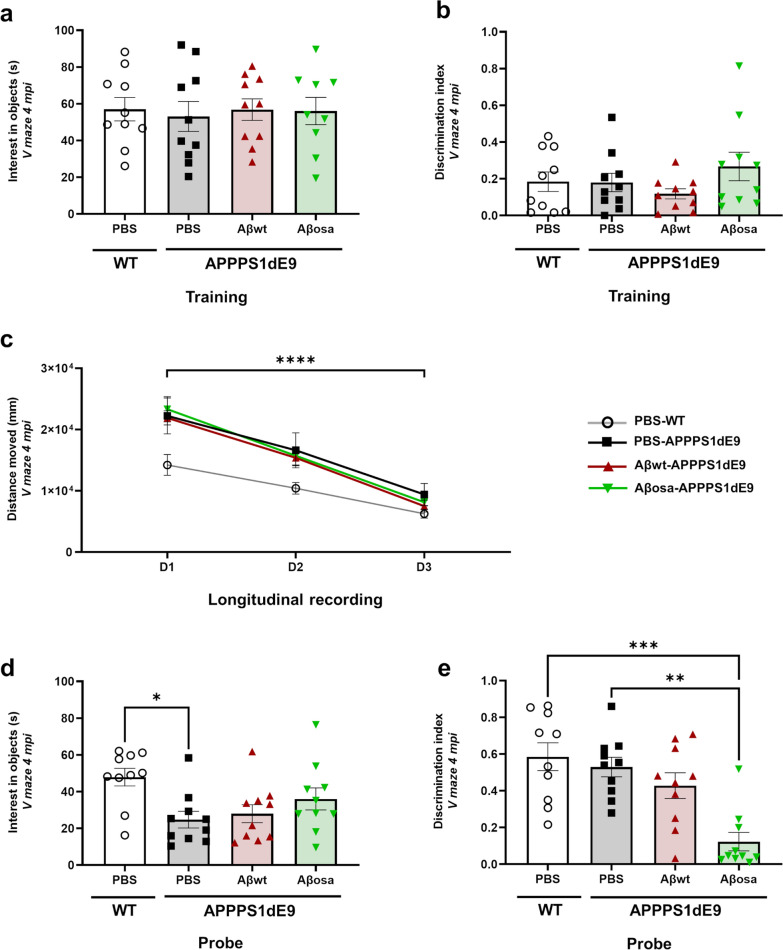
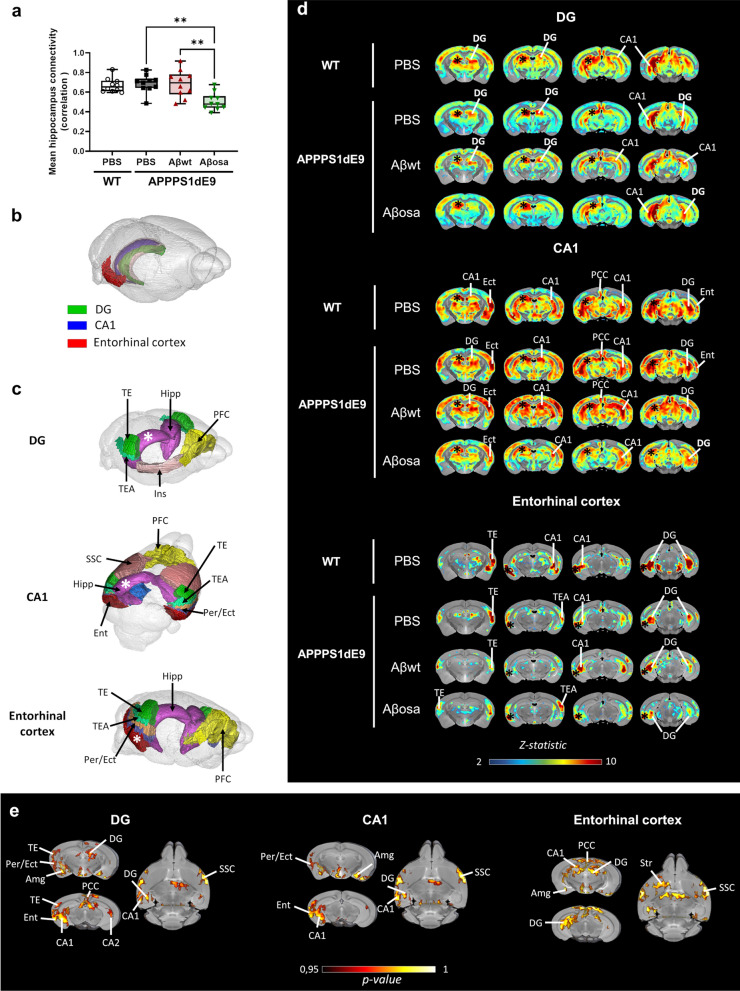
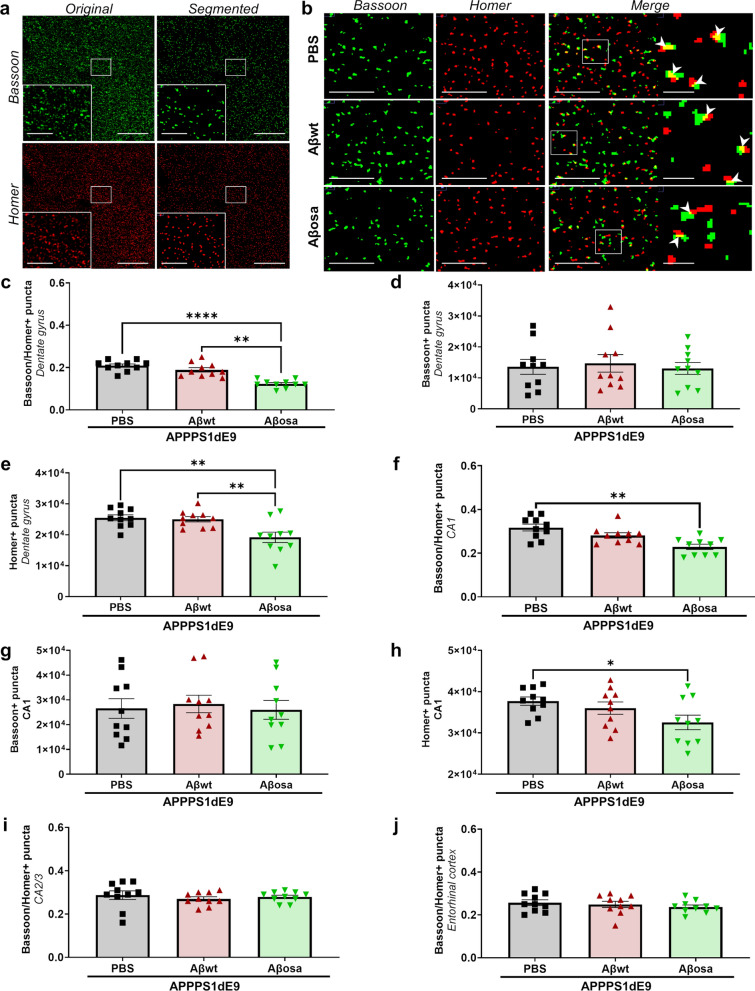
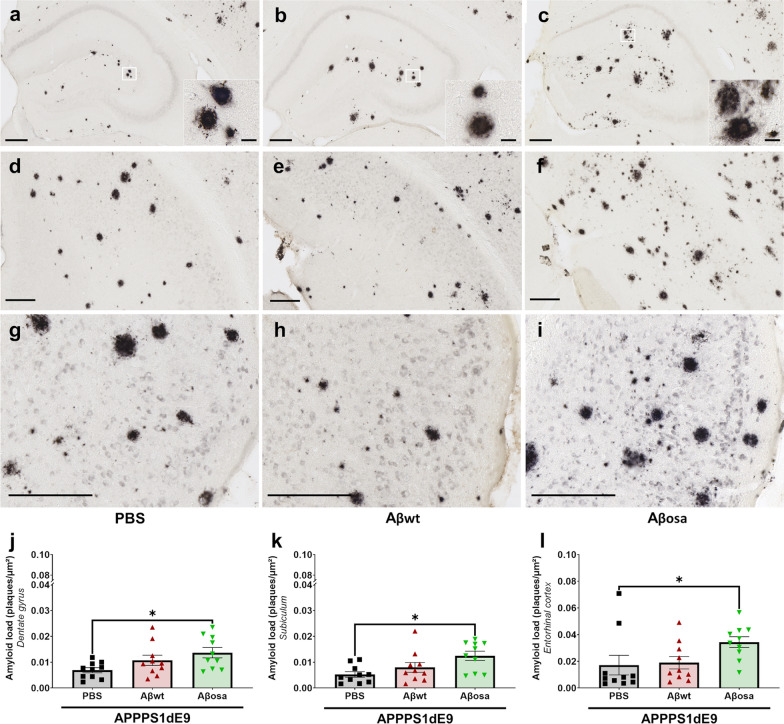
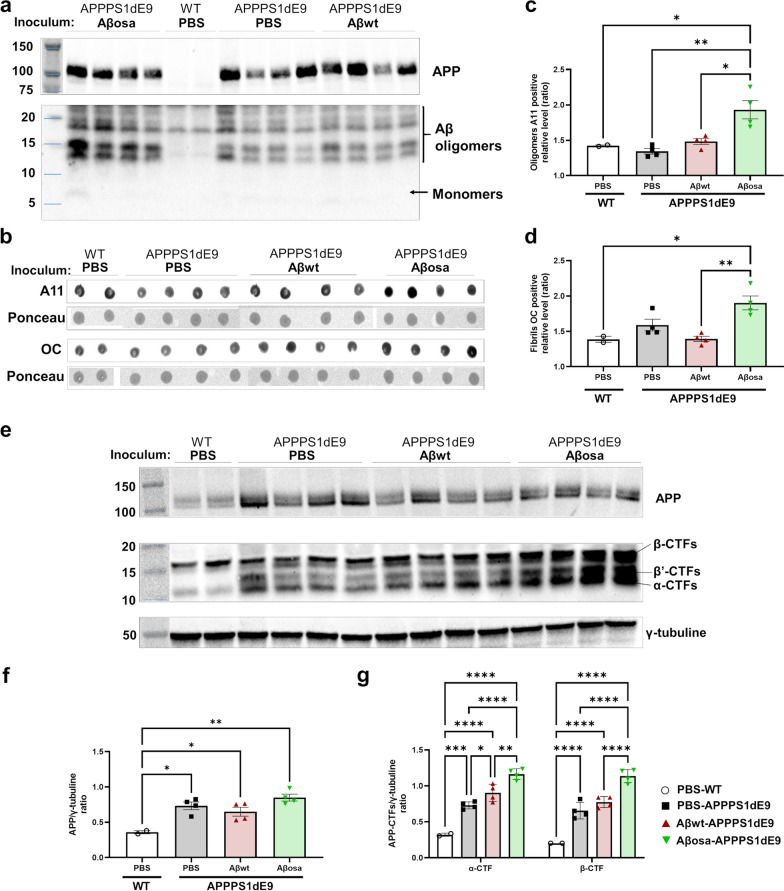
Alzheimer's disease (AD) is characterized by intracerebral deposition of abnormal proteinaceous assemblies made of amyloid-β (Aß) peptides or tau proteins. These peptides and proteins induce synaptic dysfunctions that are strongly correlated with cognitive decline. Intracerebral infusion of well-defined Aβ seeds from non-mutated Aβ1-40 or Aβ1-42 peptides can increase Aβ depositions several months after the infusion. Familial forms of AD are associated with mutations in the amyloid precursor protein (APP) that induce the production of Aβ peptides with different structures. The Aβ Osaka (Aβosa mutation (E693Δ)) is located within the Aβ sequence and thus the Aβosa peptides have different structures and properties as compared to non-mutated Aβ1-42 peptides (Aβwt). Here, we wondered if a single exposure to this mutated Aβ can worsen AD pathology as well as downstream events including cognition, cerebral connectivity and synaptic health several months after the inoculation. To answer this question we inoculated Aβ1-42-bearing Osaka mutation (Aβosa) in the dentate gyrus of APPswe/PS1dE9 mice at the age of two months. Their cognition and cerebral connectivity were analyzed at 4 months post-inoculation by behavioral evaluation and functional MRI. Aβ pathology as well as synaptic density were evaluated by histology. The impact of Aβosa peptides on synaptic health was also measured on primary cortical neurons. Remarkably, the intracerebral administration of Aβosa induced cognitive and synaptic impairments as well as a reduction of functional connectivity between different brain regions, 4 months post-inoculation. It increased Aβ plaque depositions and increased Aβ oligomers. This is the first study showing that a single, sporadic event as Aβosa inoculation can worsen the fate of the pathology and clinical outcome several months after the event. It suggests that a single inoculation of Aβ regulates a large cascade of events for a long time.
Keywords: Alzheimer’s disease; Amyloid-β; Aβ Osaka; Cerebral connectivity; Memory; Synapses.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Transmissible long-term neuroprotective and pro-cognitive effects of 1-42 beta-amyloid with A2T icelandic mutation in an Alzheimer's disease mouse model.Mol Psychiatry. 2024 Dec;29(12):3707-3721. doi: 10.1038/s41380-024-02611-8. Epub 2024 Jun 14. Mol Psychiatry. 2024. PMID: 38871852 Free PMC article.
-
Pathological changes induced by Alzheimer's brain inoculation in amyloid-beta plaque-bearing mice.Acta Neuropathol Commun. 2022 Aug 16;10(1):112. doi: 10.1186/s40478-022-01410-y. Acta Neuropathol Commun. 2022. PMID: 35974399 Free PMC article.
-
Soluble Aβ levels correlate with cognitive deficits in the 12-month-old APPswe/PS1dE9 mouse model of Alzheimer's disease.Behav Brain Res. 2011 Sep 23;222(2):342-50. doi: 10.1016/j.bbr.2011.03.072. Epub 2011 Apr 14. Behav Brain Res. 2011. PMID: 21513747
-
APP transgenic modeling of Alzheimer's disease: mechanisms of neurodegeneration and aberrant neurogenesis.Brain Struct Funct. 2010 Mar;214(2-3):111-26. doi: 10.1007/s00429-009-0232-6. Epub 2009 Nov 29. Brain Struct Funct. 2010. PMID: 20091183 Free PMC article. Review.
-
APP Osaka Mutation in Familial Alzheimer's Disease-Its Discovery, Phenotypes, and Mechanism of Recessive Inheritance.Int J Mol Sci. 2020 Feb 19;21(4):1413. doi: 10.3390/ijms21041413. Int J Mol Sci. 2020. PMID: 32093100 Free PMC article. Review.
Cited by
-
Transmissible long-term neuroprotective and pro-cognitive effects of 1-42 beta-amyloid with A2T icelandic mutation in an Alzheimer's disease mouse model.Mol Psychiatry. 2024 Dec;29(12):3707-3721. doi: 10.1038/s41380-024-02611-8. Epub 2024 Jun 14. Mol Psychiatry. 2024. PMID: 38871852 Free PMC article.
-
Cognitive integrity in Non-Demented Individuals with Alzheimer's Neuropathology is associated with preservation and remodeling of dendritic spines.Alzheimers Dement. 2024 Jul;20(7):4677-4691. doi: 10.1002/alz.13900. Epub 2024 Jun 3. Alzheimers Dement. 2024. PMID: 38829680 Free PMC article.
-
In Vivo Seeding of Amyloid-β Protein and Implications in Modeling Alzheimer's Disease Pathology.Biomolecules. 2025 Apr 11;15(4):571. doi: 10.3390/biom15040571. Biomolecules. 2025. PMID: 40305318 Free PMC article. Review.
References
-
- Balducci C, Beeg M, Stravalaci M, Bastone A, Sclip A, Biasini E, Tapella L, Colombo L, Manzoni C, Borsello T, Chiesa R, Gobbi M, Salmona M, Forloni G. Synthetic amyloid-beta oligomers impair long-term memory independently of cellular prion protein. Proc Natl Acad Sci USA. 2010;107:2295–2300. doi: 10.1073/pnas.0911829107. - DOI - PMC - PubMed
-
- du Sert NP, Hurst V, Ahluwalia A, Alam S, Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl U, Emerson M, Garner P, Holgate ST, Howells DW, Karp NA, Lazic SE, Lidster K, MacCallum CJ, Macleod M, Pearl EJ, Petersen OH, Rawle F, Reynolds P, Rooney K, Sena ES, Silberberg SD, Steckler T, Wurbel H. The ARRIVE guidelines 20: Updated guidelines for reporting animal research. Bmc Veterinary Research. 2020 doi: 10.1186/s12917-020-02451-y. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
