

The ferroptosis inducing compounds RSL3 and ML162 are not direct inhibitors of GPX4 but of TXNRD1
- PMID: 37087975
- PMCID: PMC10149367
- DOI: 10.1016/j.redox.2023.102703
The ferroptosis inducing compounds RSL3 and ML162 are not direct inhibitors of GPX4 but of TXNRD1
Abstract
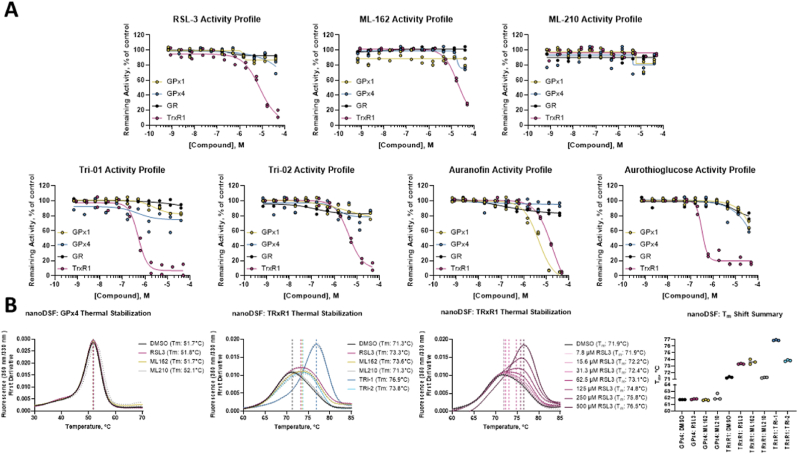
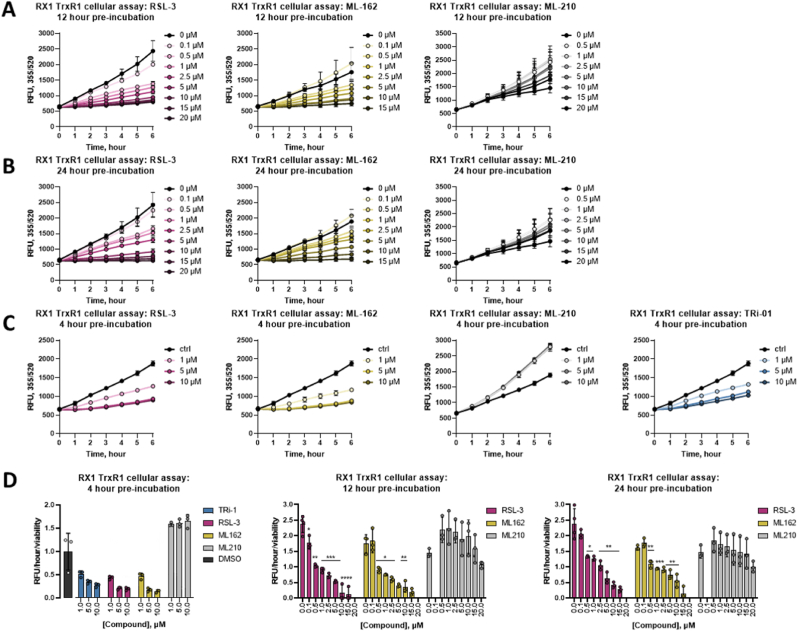
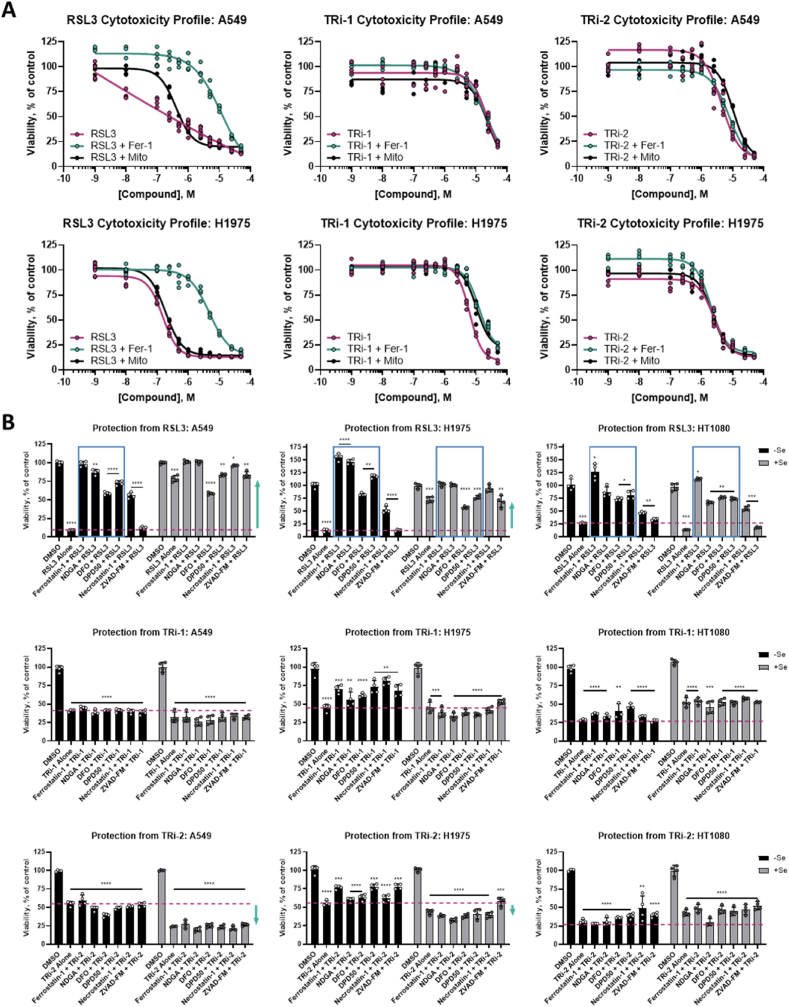
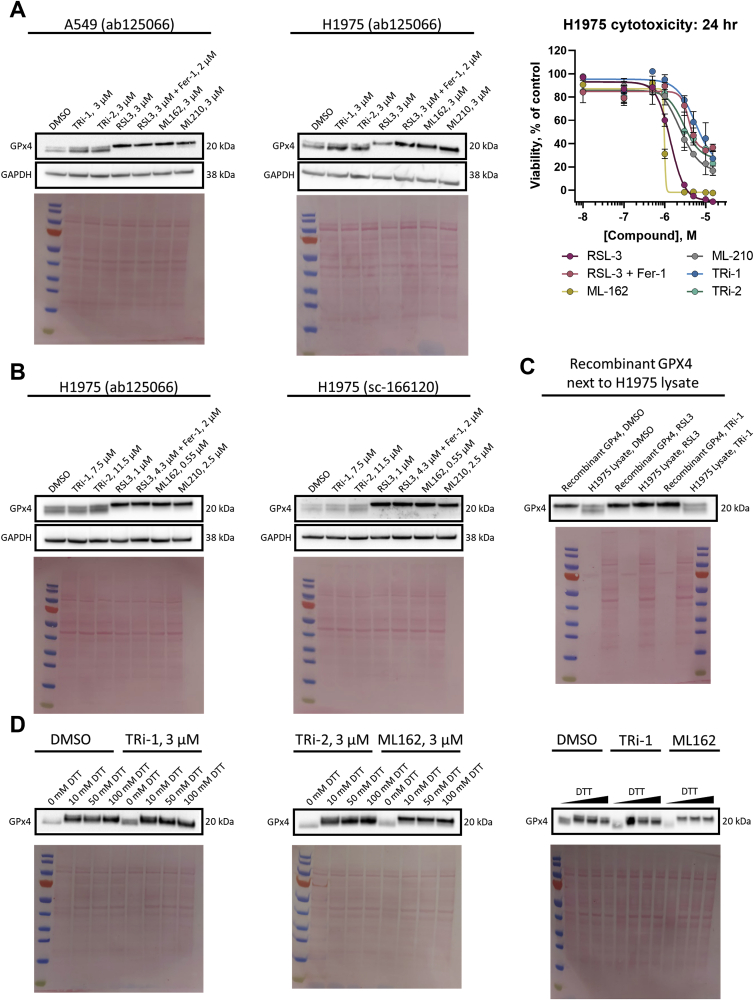
Ferroptosis is defined as cell death triggered by iron-dependent lipid peroxidation that is preventable by antioxidant compounds such as ferrostatin-1. Endogenous suppressors of ferroptosis include FSP-1 and the selenoprotein GPX4, the latter of which directly enzymatically reduces lipid hydroperoxides. Small molecules that trigger ferroptosis include RSL3, ML162, and ML210; these compounds are often used in studies of ferroptosis and are generally considered as GPX4 inhibitors. Here, we found that RSL3 and ML162 completely lack capacity of inhibiting the enzymatic activity of recombinant selenoprotein GPX4. Surprisingly, these compounds were instead found to be efficient inhibitors of another selenoprotein, TXNRD1. Other known inhibitors of TXNRD1, including auranofin, TRi-1 and TRi-2, are also efficient inducers of cell death but that cell death could not be suppressed with ferrostatin-1. Our results collectively suggest that prior studies using RSL3 and ML162 may need to be reevaluated in the context of ferroptosis with regards to additional enzyme targets and mechanisms of action that may be involved.
Copyright © 2023 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest ESJA has patents on TRi compounds for development towards cancer treatment. ESJA and QC are shareholders in Selenozyme AB, a company selling recombinant selenoproteins.
Figures
Similar articles
-
Lipid Peroxidation-Dependent Cell Death Regulated by GPx4 and Ferroptosis.Curr Top Microbiol Immunol. 2017;403:143-170. doi: 10.1007/82_2016_508. Curr Top Microbiol Immunol. 2017. PMID: 28204974 Review.
-
Discovery of Novel Potent Covalent Glutathione Peroxidase 4 Inhibitors as Highly Selective Ferroptosis Inducers for the Treatment of Triple-Negative Breast Cancer.J Med Chem. 2023 Jul 27;66(14):10036-10059. doi: 10.1021/acs.jmedchem.3c00967. Epub 2023 Jul 15. J Med Chem. 2023. PMID: 37452764
-
Slowly progressive cell death induced by GPx4-deficiency occurs via MEK1/ERK2 activation as a downstream signal after iron-independent lipid peroxidation.J Clin Biochem Nutr. 2024 Mar;74(2):97-107. doi: 10.3164/jcbn.23-101. Epub 2023 Nov 1. J Clin Biochem Nutr. 2024. PMID: 38510679 Free PMC article.
-
Mitochondrial rescue prevents glutathione peroxidase-dependent ferroptosis.Free Radic Biol Med. 2018 Mar;117:45-57. doi: 10.1016/j.freeradbiomed.2018.01.019. Epub 2018 Jan 31. Free Radic Biol Med. 2018. PMID: 29378335
-
A white paper on Phospholipid Hydroperoxide Glutathione Peroxidase (GPx4) forty years later.Free Radic Biol Med. 2022 Aug 1;188:117-133. doi: 10.1016/j.freeradbiomed.2022.06.227. Epub 2022 Jun 16. Free Radic Biol Med. 2022. PMID: 35718302 Review.
Cited by
-
Insights into Free Drug Release from Efficacious N-Acyl O-Aminophenol Duocarmycin Prodrugs.ACS Chem Biol. 2025 Feb 21;20(2):442-454. doi: 10.1021/acschembio.4c00754. Epub 2025 Feb 9. ACS Chem Biol. 2025. PMID: 39924956
-
The crosstalk between glutathione metabolism and non-coding RNAs in cancer progression and treatment resistance.Redox Biol. 2025 Jul;84:103689. doi: 10.1016/j.redox.2025.103689. Epub 2025 May 19. Redox Biol. 2025. PMID: 40403492 Free PMC article. Review.
-
Ferroptosis and Metabolic Dysregulation: Emerging Chemical Targets in Cancer and Infection.Molecules. 2025 Jul 18;30(14):3020. doi: 10.3390/molecules30143020. Molecules. 2025. PMID: 40733290 Free PMC article. Review.
-
Recharacterization of the Tumor Suppressive Mechanism of RSL3 Identifies the Selenoproteome as a Druggable Pathway in Colorectal Cancer.Cancer Res. 2025 Aug 1;85(15):2788-2804. doi: 10.1158/0008-5472.CAN-24-3478. Cancer Res. 2025. PMID: 40392234
-
Ironing out the details: ferroptosis and its relevance to diabetic cardiomyopathy.Am J Physiol Regul Integr Comp Physiol. 2023 Dec 1;325(6):R665-R681. doi: 10.1152/ajpregu.00117.2023. Epub 2023 Sep 25. Am J Physiol Regul Integr Comp Physiol. 2023. PMID: 37746707 Free PMC article. Review.
References
-
- Maiorino M., Conrad M., Ursini F. GPx4, lipid peroxidation, and cell death: discoveries, rediscoveries, and open issues. Antioxidants Redox Signal. 2018;29:61–74. - PubMed
-
- Viswanathan V.S., Ryan M.J., Dhruv H.D., Gill S., Eichhoff O.M., Seashore-Ludlow B., Kaffenberger S.D., Eaton J.K., Shimada K., Aguirre A.J., Viswanathan S.R., Chattopadhyay S., Tamayo P., Yang W.S., Rees M.G., Chen S., Boskovic Z.V., Javaid S., Huang C., Wu X., Tseng Y.Y., Roider E.M., Gao D., Cleary J.M., Wolpin B.M., Mesirov J.P., Haber D.A., Engelman J.A., Boehm J.S., Kotz J.D., Hon C.S., Chen Y., Hahn W.C., Levesque M.P., Doench J.G., Berens M.E., Shamji A.F., Clemons P.A., Stockwell B.R., Schreiber S.L. Dependency of a therapy-resistant state of cancer cells on a lipid peroxidase pathway. Nature. 2017;547:453–457. - PMC - PubMed
-
- Ursini F., Maiorino M. Lipid peroxidation and ferroptosis: the role of GSH and GPx4. Free Radic. Biol. Med. 2020;152:175–185. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
