

This is a preprint.
Rbm8a deficiency causes hematopoietic defects by modulating Wnt/PCP signaling
- PMID: 37090609
- PMCID: PMC10120739
- DOI: 10.1101/2023.04.12.536513
Rbm8a deficiency causes hematopoietic defects by modulating Wnt/PCP signaling
Update in
-
Rbm8a deficiency causes hematopoietic defects by modulating Wnt/PCP signaling.Dev Biol. 2025 Dec;528:34-56. doi: 10.1016/j.ydbio.2025.08.021. Epub 2025 Sep 2. Dev Biol. 2025. PMID: 40907933 Free PMC article.
Abstract
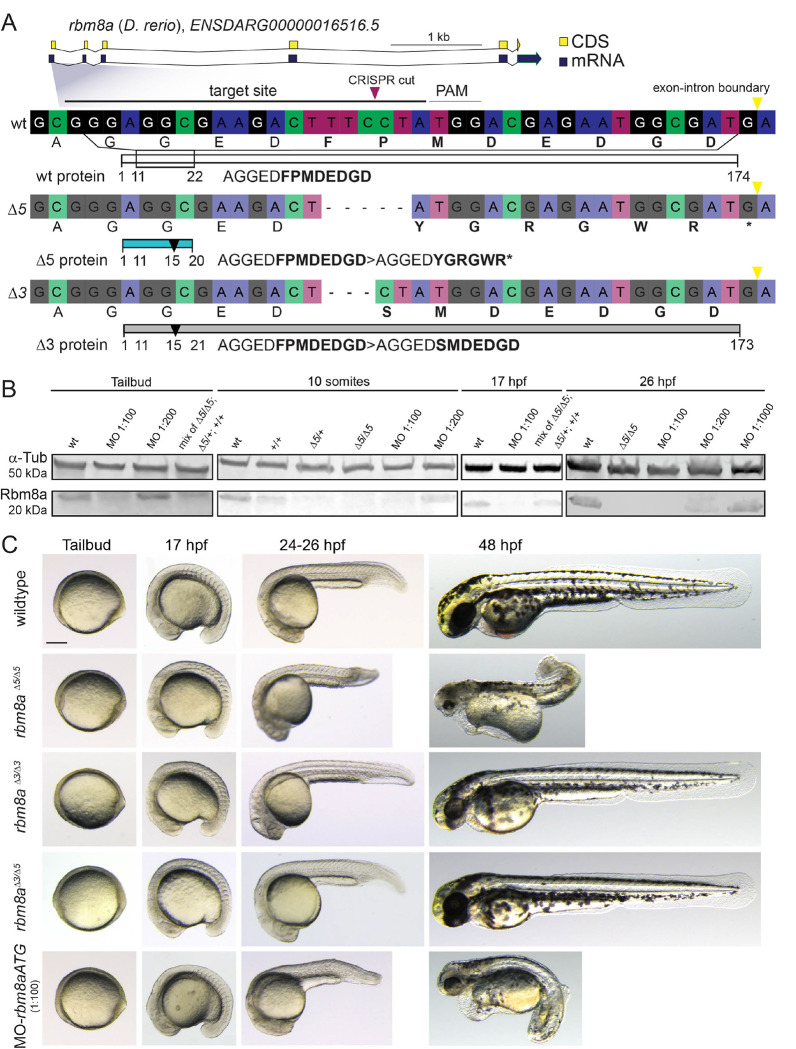
Defects in blood development frequently occur among syndromic congenital anomalies. Thrombocytopenia-Absent Radius (TAR) syndrome is a rare congenital condition with reduced platelets (hypomegakaryocytic thrombocytopenia) and forelimb anomalies, concurrent with more variable heart and kidney defects. TAR syndrome associates with hypomorphic gene function for RBM8A/Y14 that encodes a component of the exon junction complex involved in mRNA splicing, transport, and nonsense-mediated decay. How perturbing a general mRNA-processing factor causes the selective TAR Syndrome phenotypes remains unknown. Here, we connect zebrafish rbm8a perturbation to early hematopoietic defects via attenuated non-canonical Wnt/Planar Cell Polarity (PCP) signaling that controls developmental cell re-arrangements. In hypomorphic rbm8a zebrafish, we observe a significant reduction of cd41-positive thrombocytes. rbm8a-mutant zebrafish embryos accumulate mRNAs with individual retained introns, a hallmark of defective nonsense-mediated decay; affected mRNAs include transcripts for non-canonical Wnt/PCP pathway components. We establish that rbm8a-mutant embryos show convergent extension defects and that reduced rbm8a function interacts with perturbations in non-canonical Wnt/PCP pathway genes wnt5b, wnt11f2, fzd7a, and vangl2. Using live-imaging, we found reduced rbm8a function impairs the architecture of the lateral plate mesoderm (LPM) that forms hematopoietic, cardiovascular, kidney, and forelimb skeleton progenitors as affected in TAR Syndrome. Both mutants for rbm8a and for the PCP gene vangl2 feature impaired expression of early hematopoietic/endothelial genes including runx1 and the megakaryocyte regulator gfi1aa. Together, our data propose aberrant LPM patterning and hematopoietic defects as consequence of attenuated non-canonical Wnt/PCP signaling upon reduced rbm8a function. These results also link TAR Syndrome to a potential LPM origin and a developmental mechanism.
Keywords: development; hematopoiesis; morphogenesis; non-canonical Wnt; thrombocytopenia; zebrafish.
Conflict of interest statement
COMPETING INTERESTS STATEMENT J.A.S. has been a consultant for Sanofi, Takeda, Genentech, CSL Behring, and HEMA Biologics.
Figures






References
-
- Petit F. & Boussion S. Thrombocytopenia Absent Radius Syndrome. GeneReviews® (2022). - PubMed
-
- Albers C. A., Newbury-Ecob R., Ouwehand W. H. & Ghevaert C. New insights into the genetic basis of TAR (thrombocytopenia-absent radii) syndrome. Curr Opin Genet Dev 23, 316–23 (2013). - PubMed
-
- Boussion S. et al. TAR syndrome: clinical and molecular characterization of a cohort of 26 patients and description of novel non-coding variants of RBM8A. Hum Mutat 41, 1220–1225 (2020). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources