

This is a preprint.
FGF-21 Conducts a Liver-Brain-Kidney Axis to Promote Renal Cell Carcinoma
- PMID: 37090652
- PMCID: PMC10120688
- DOI: 10.1101/2023.04.12.536558
FGF-21 Conducts a Liver-Brain-Kidney Axis to Promote Renal Cell Carcinoma
Abstract
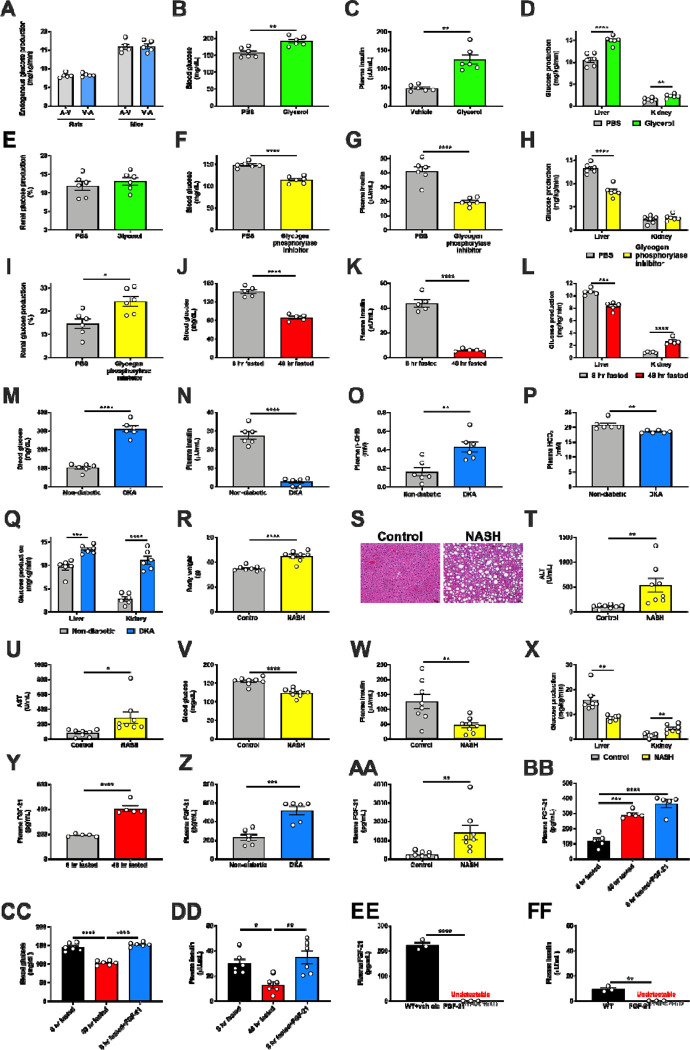
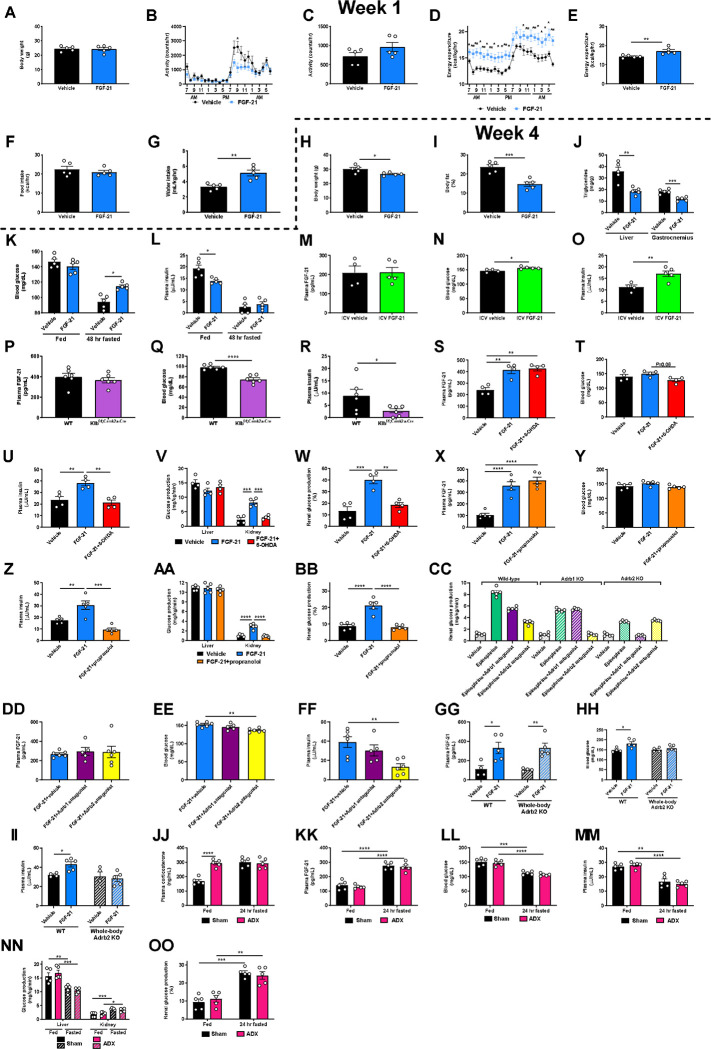
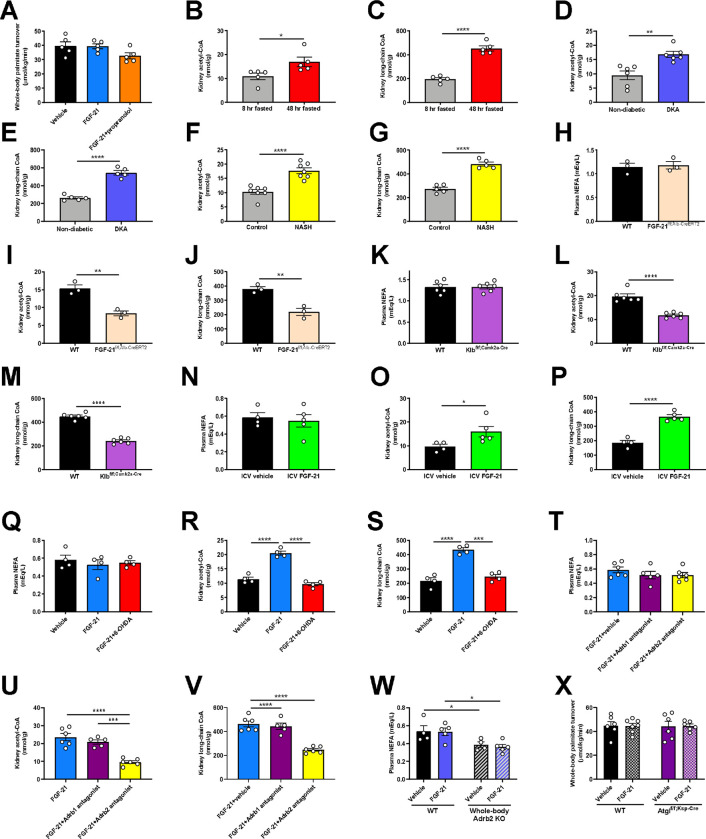
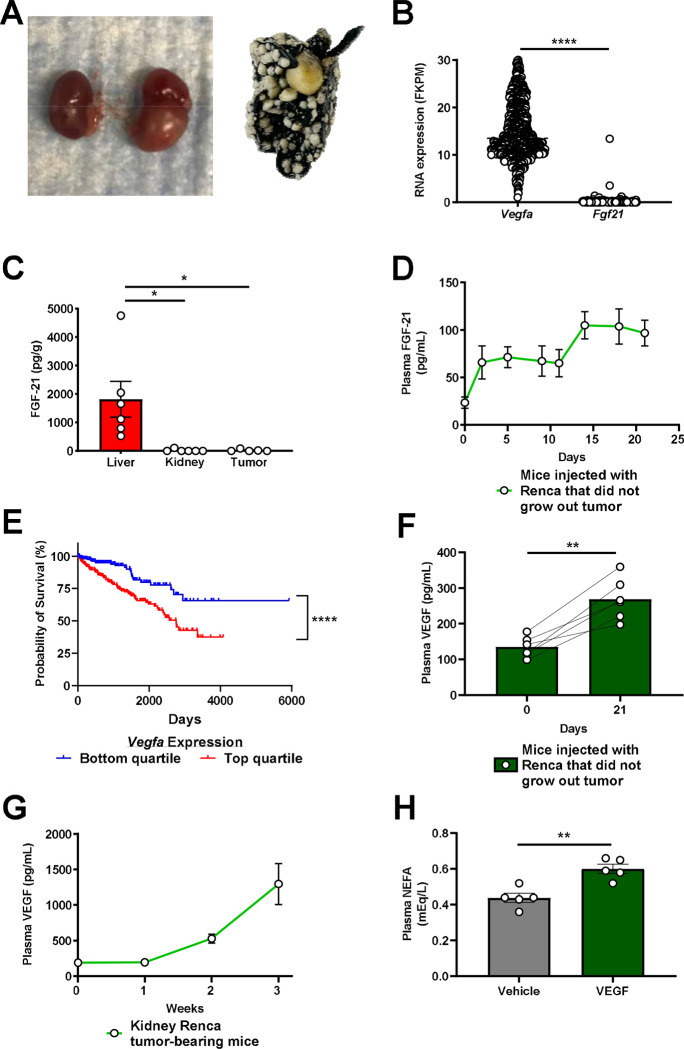
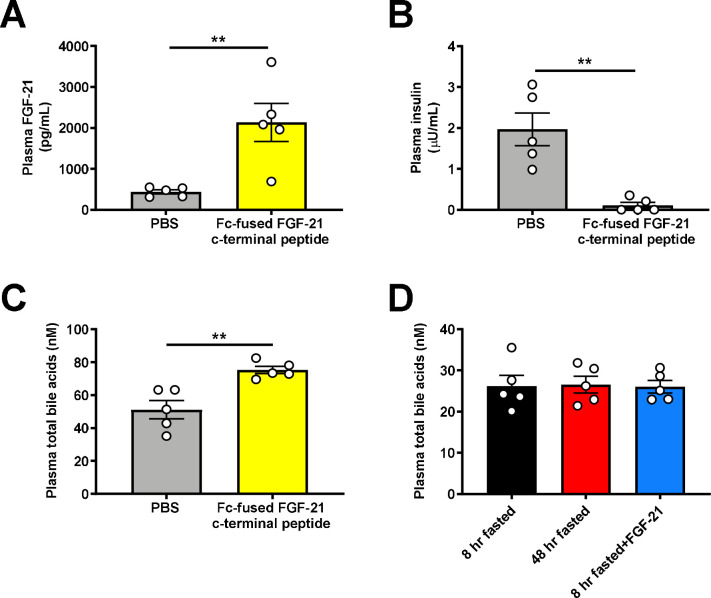
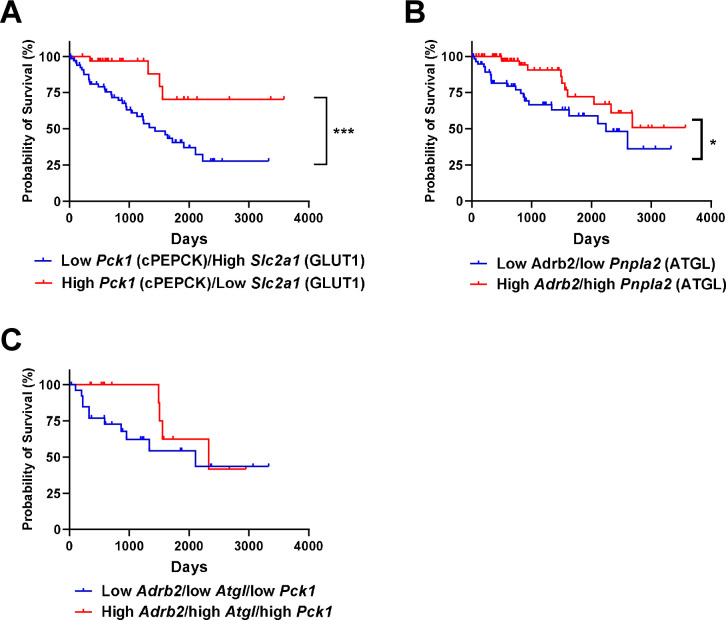
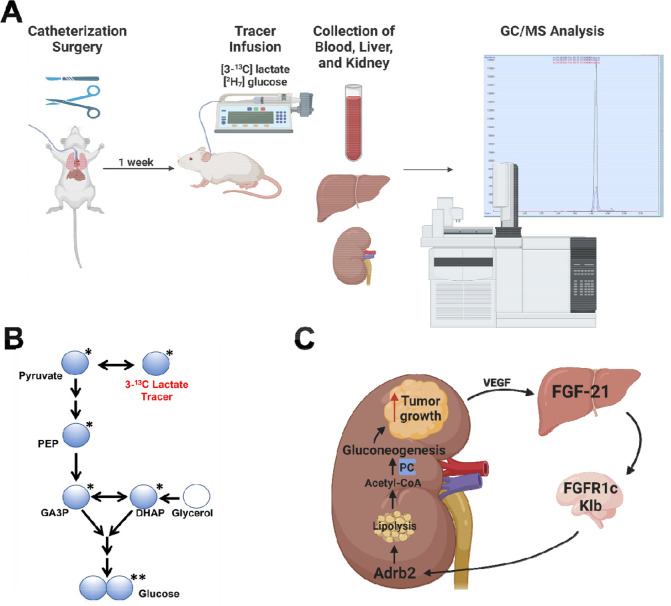
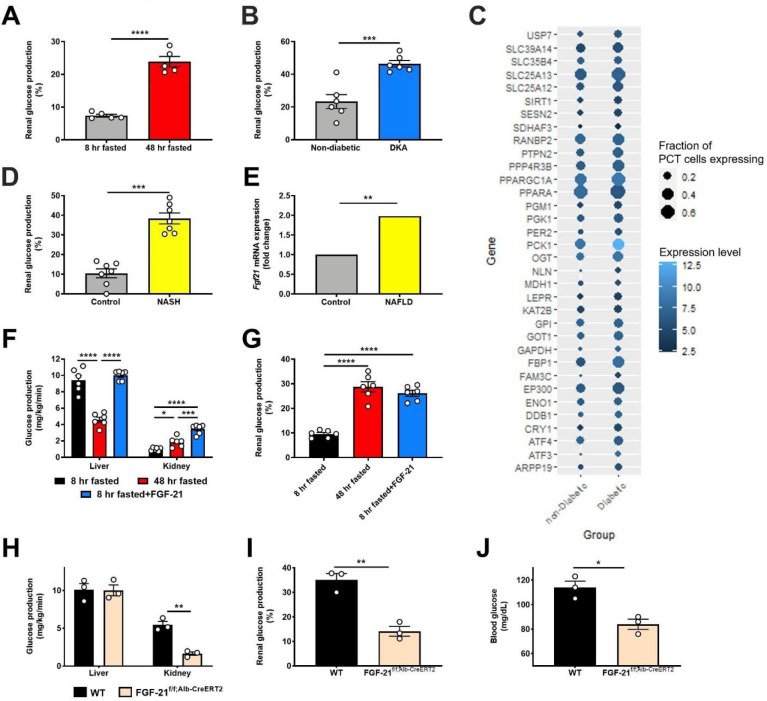
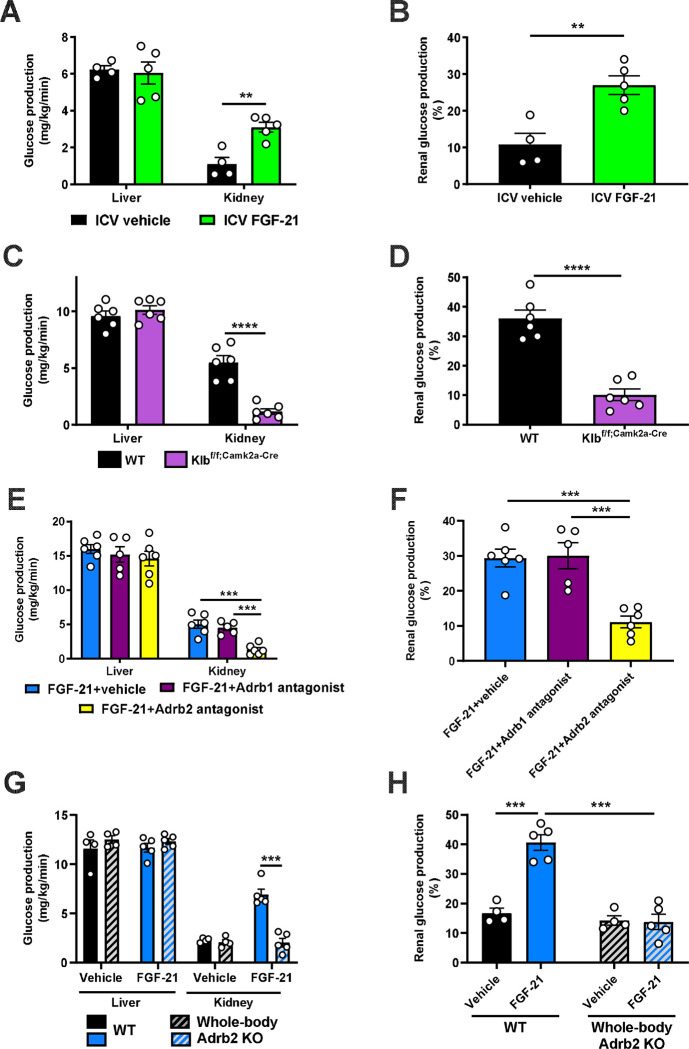
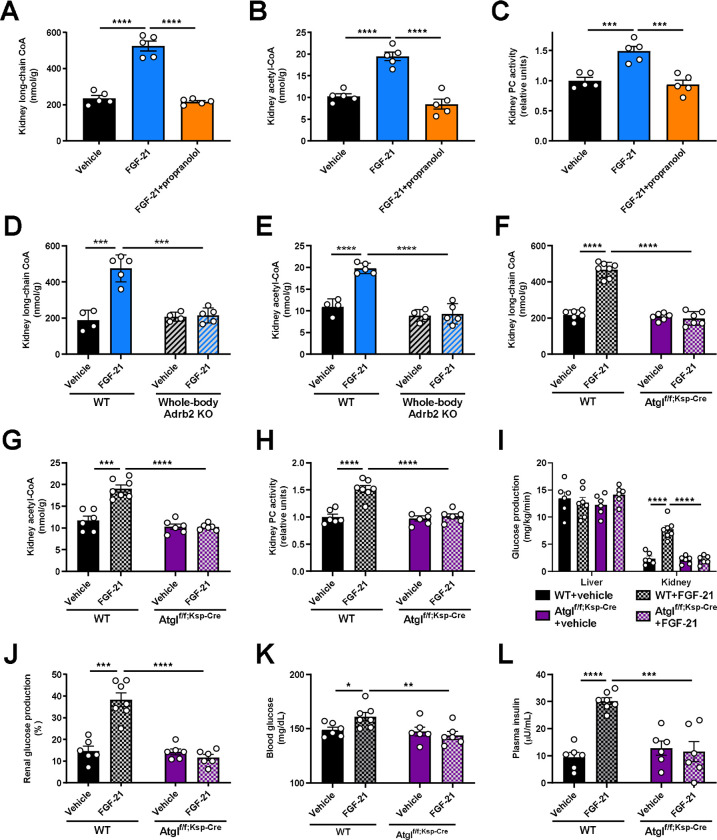
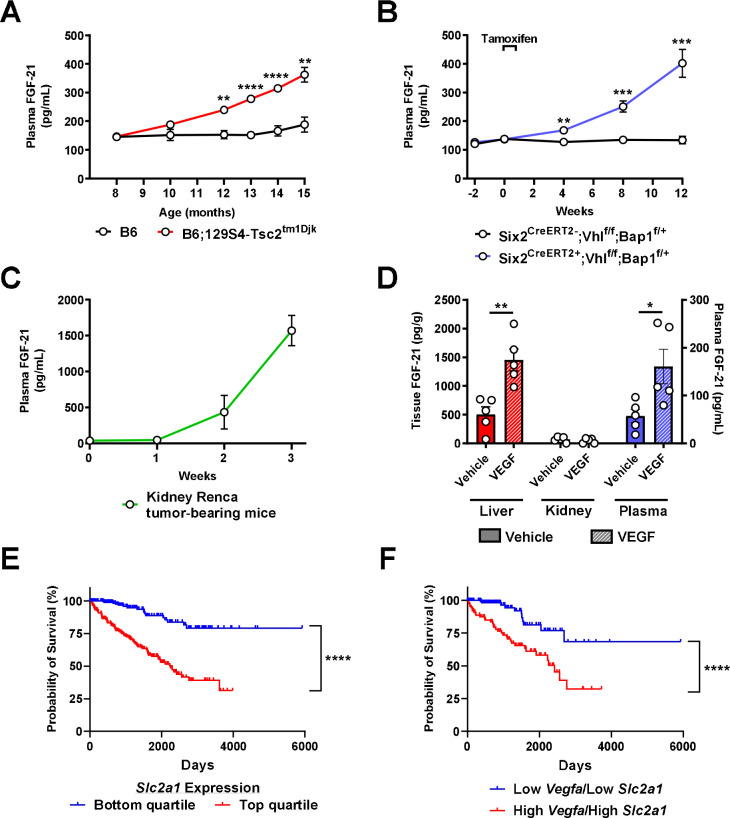
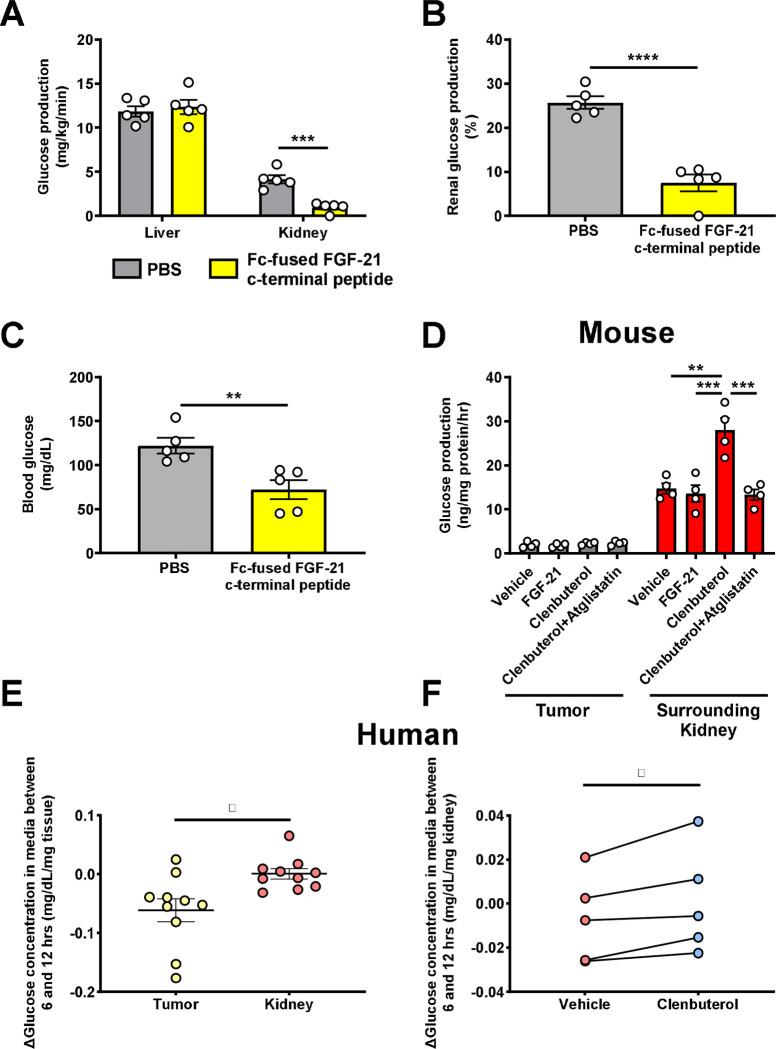
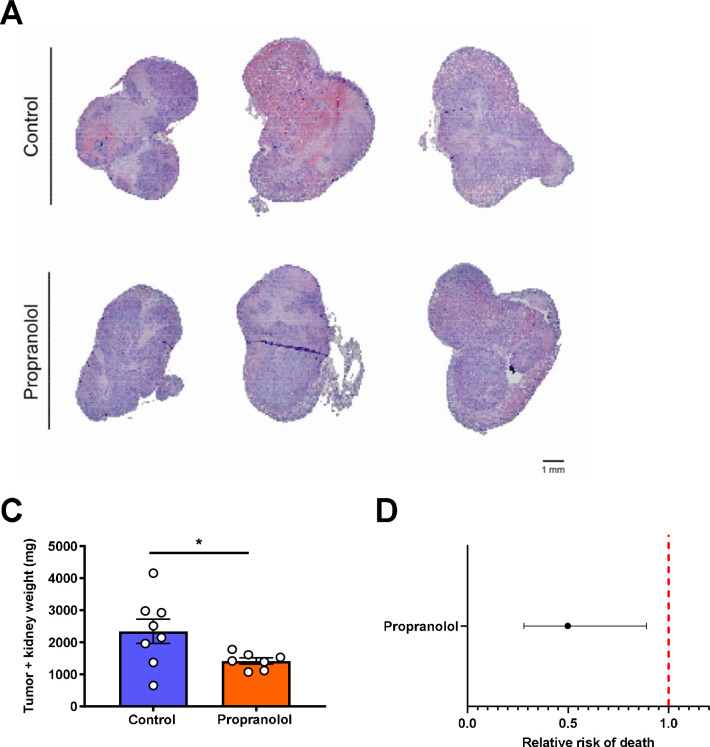
Metabolic homeostasis is one of the most exquisitely tuned systems in mammalian physiology. Metabolic homeostasis requires multiple redundant systems to cooperate to maintain blood glucose concentrations in a narrow range, despite a multitude of physiological and pathophysiological pressures. Cancer is one of the canonical pathophysiological settings in which metabolism plays a key role. In this study, we utilized REnal Gluconeogenesis Analytical Leads (REGAL), a liquid chromatography-mass spectrometry/mass spectrometry-based stable isotope tracer method that we developed to show that in conditions of metabolic stress, the fasting hepatokine fibroblast growth factor-21 (FGF-21)1,2 coordinates a liver-brain-kidney axis to promote renal gluconeogenesis. FGF-21 promotes renal gluconeogenesis by enhancing β2 adrenergic receptor (Adrb2)-driven, adipose triglyceride lipase (ATGL)-mediated intrarenal lipolysis. Further, we show that this liver-brain-kidney axis promotes gluconeogenesis in the renal parenchyma in mice and humans with renal cell carcinoma (RCC). This increased gluconeogenesis is, in turn, associated with accelerated RCC progression. We identify Adrb2 blockade as a new class of therapy for RCC in mice, with confirmatory data in human patients. In summary, these data reveal a new metabolic function of FGF-21 in driving renal gluconeogenesis, and demonstrate that inhibition of renal gluconeogenesis by FGF-21 antagonism deserves attention as a new therapeutic approach to RCC.
Conflict of interest statement
Competing Interests Declaration The authors declare no competing interests.
Figures
References
-
- Badman M. K. et al. Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab 5, 426–437 (2007). - PubMed
-
- Inagaki T. et al. Endocrine regulation of the fasting response by PPARalpha-mediated induction of fibroblast growth factor 21. Cell Metab 5, 415–425 (2007). - PubMed
-
- Renehan A. G., Tyson M., Egger M., Heller R. F. & Zwahlen M. Body-mass index and incidence of cancer: a systematic review and meta-analysis of prospective observational studies. Lancet 371, 569–578 (2008). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources