

Regulation of High-Affinity Iron Acquisition, Including Acquisition Mediated by the Iron Permease FtrA, Is Coordinated by AtrR, SrbA, and SreA in Aspergillus fumigatus
- PMID: 37093084
- PMCID: PMC10294635
- DOI: 10.1128/mbio.00757-23
Regulation of High-Affinity Iron Acquisition, Including Acquisition Mediated by the Iron Permease FtrA, Is Coordinated by AtrR, SrbA, and SreA in Aspergillus fumigatus
Abstract
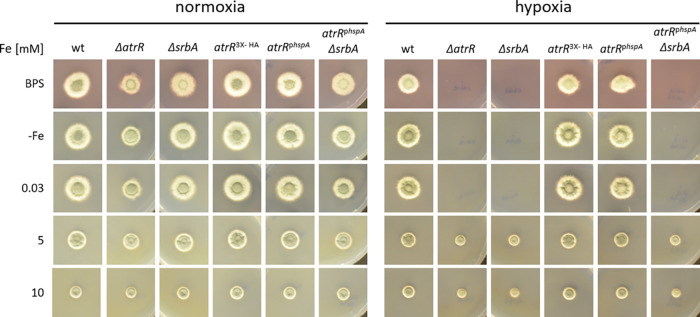
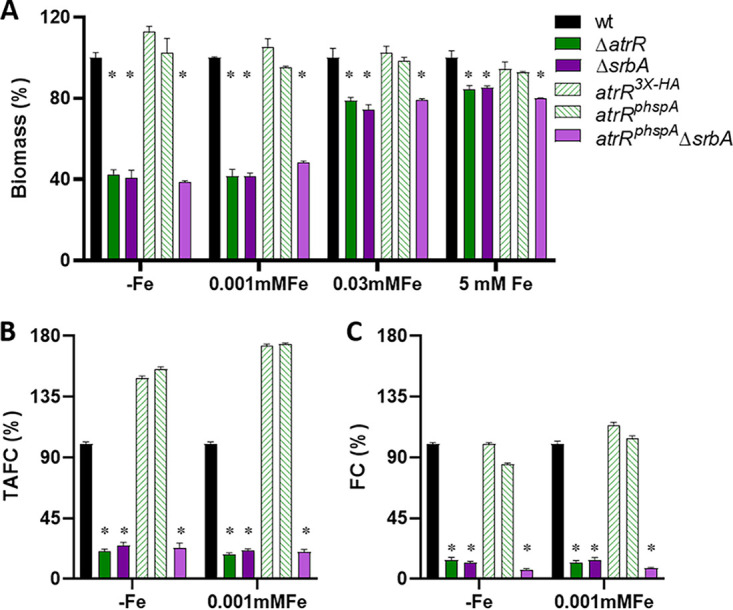
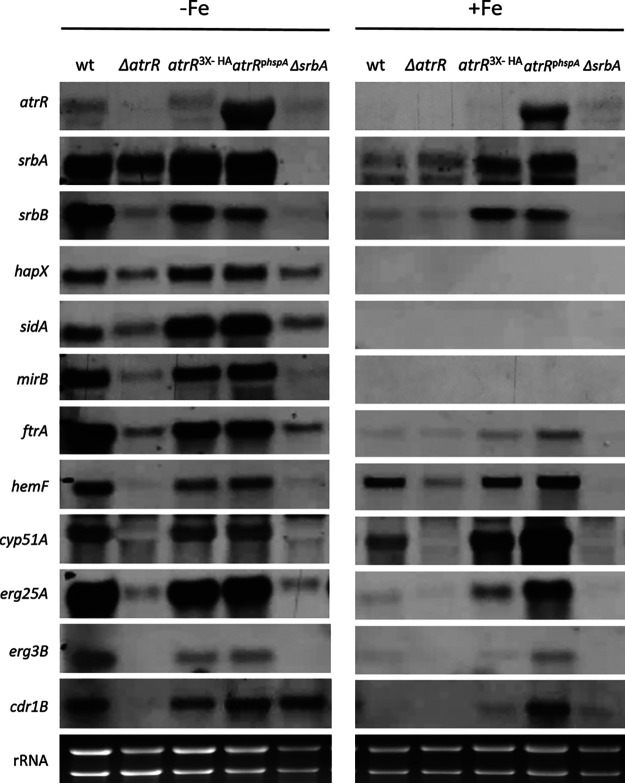
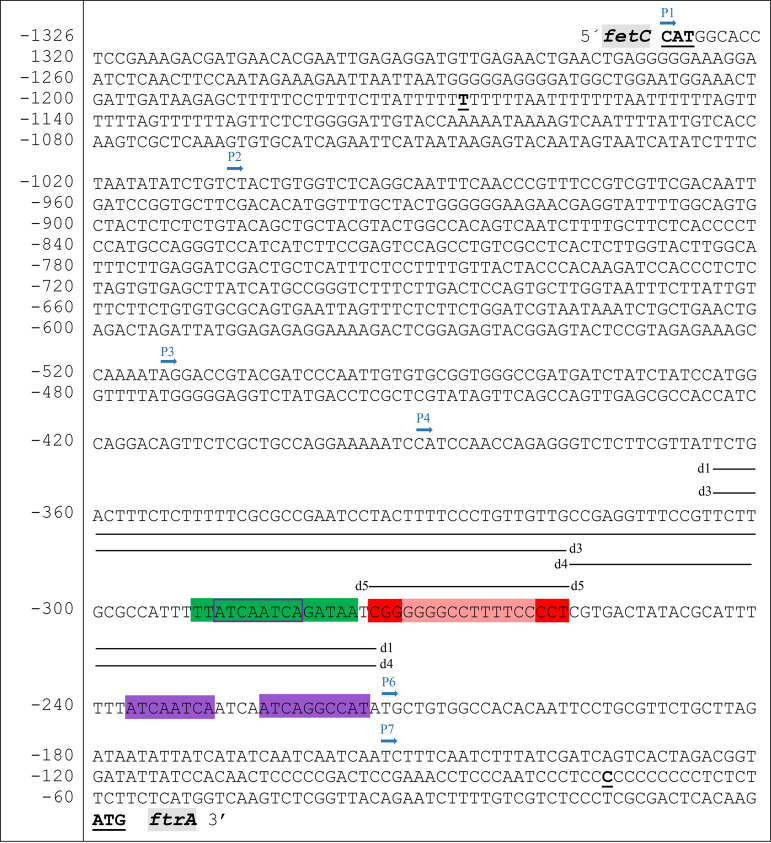
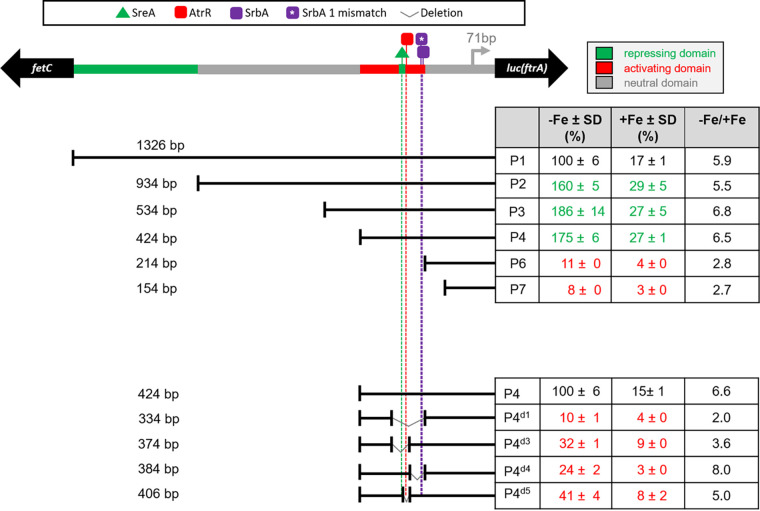
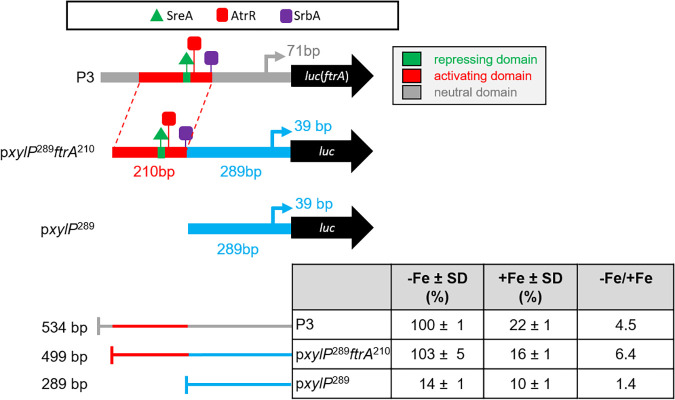
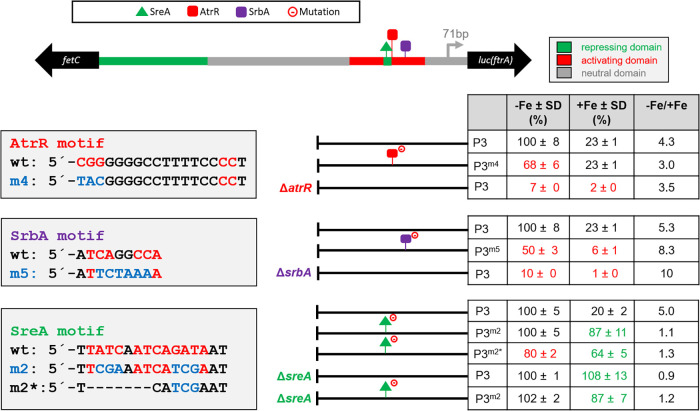
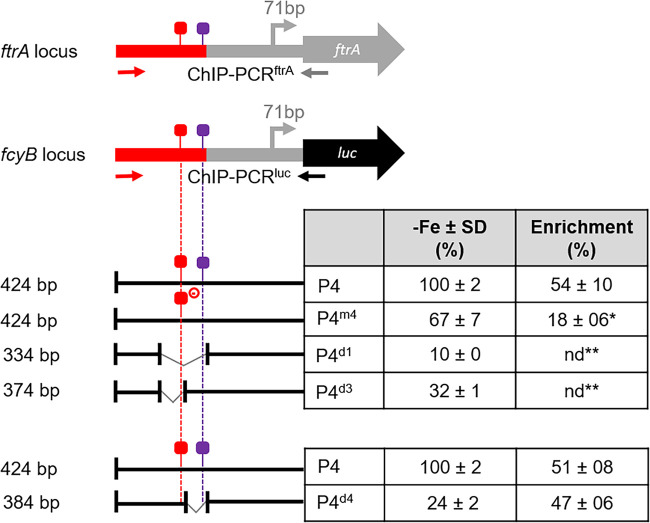
Iron acquisition is crucial for virulence of the human pathogen Aspergillus fumigatus. Previous studies indicated that this mold regulates iron uptake via both siderophores and reductive iron assimilation by the GATA factor SreA and the SREBP regulator SrbA. Here, characterization of loss of function as well as hyperactive alleles revealed that transcriptional activation of iron uptake depends additionally on the Zn2Cys6 regulator AtrR, most likely via cooperation with SrbA. Mutational analysis of the promoter of the iron permease-encoding ftrA gene identified a 210-bp sequence, which is both essential and sufficient to impart iron regulation. Further studies located functional sequences, densely packed within 75 bp, that largely resemble binding motifs for SrbA, SreA, and AtrR. The latter, confirmed by chromatin immunoprecipitation (ChIP) analysis, is the first one not fully matching the 5'-CGGN12CCG-3' consensus sequence. The results presented here emphasize for the first time the direct involvement of SrbA, AtrR, and SreA in iron regulation. The essential role of both AtrR and SrbA in activation of iron acquisition underlines the coordination of iron homeostasis with biosynthesis of ergosterol and heme as well as adaptation to hypoxia. The rationale is most likely the iron dependence of these pathways along with the enzymatic link of biosynthesis of ergosterol and siderophores. IMPORTANCE Aspergillus fumigatus is the most common filamentous fungal pathogen infecting humans. Iron acquisition via siderophores has previously been shown to be essential for virulence of this mold species. Here, we demonstrate that AtrR, a transcription factor previously shown to control ergosterol biosynthesis, azole resistance, and adaptation to hypoxia, is essential for activation of iron acquisition, including siderophore biosynthesis and uptake. Dissection of an iron-regulated promoter identified binding motifs for AtrR and the two previously identified regulators of iron acquisition, SrbA and SreA. Altogether, this study identified a new regulator required for maintenance of iron homeostasis, revealed insights into promoter architecture for iron regulation, and emphasized the coordinated regulation of iron homeostasis ergosterol biosynthesis and adaptation to hypoxia.
Keywords: Aspergillus fumigatus; fungi; iron; molds; regulation; siderophore; siderophores; transcription factor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
SREBP coordinates iron and ergosterol homeostasis to mediate triazole drug and hypoxia responses in the human fungal pathogen Aspergillus fumigatus.PLoS Genet. 2011 Dec;7(12):e1002374. doi: 10.1371/journal.pgen.1002374. Epub 2011 Dec 1. PLoS Genet. 2011. PMID: 22144905 Free PMC article.
-
SreA-mediated iron regulation in Aspergillus fumigatus.Mol Microbiol. 2008 Oct;70(1):27-43. doi: 10.1111/j.1365-2958.2008.06376.x. Epub 2008 Aug 21. Mol Microbiol. 2008. PMID: 18721228 Free PMC article.
-
ChIP-seq and in vivo transcriptome analyses of the Aspergillus fumigatus SREBP SrbA reveals a new regulator of the fungal hypoxia response and virulence.PLoS Pathog. 2014 Nov 6;10(11):e1004487. doi: 10.1371/journal.ppat.1004487. eCollection 2014 Nov. PLoS Pathog. 2014. PMID: 25375670 Free PMC article.
-
Fungal iron homeostasis with a focus on Aspergillus fumigatus.Biochim Biophys Acta Mol Cell Res. 2021 Jan;1868(1):118885. doi: 10.1016/j.bbamcr.2020.118885. Epub 2020 Oct 10. Biochim Biophys Acta Mol Cell Res. 2021. PMID: 33045305 Review.
-
The crucial role of iron uptake in Aspergillus fumigatus virulence.Curr Opin Microbiol. 2013 Dec;16(6):692-9. doi: 10.1016/j.mib.2013.07.012. Epub 2013 Aug 17. Curr Opin Microbiol. 2013. PMID: 23962820 Review.
Cited by
-
Exploring the Siderophore Portfolio for Mass Spectrometry-Based Diagnosis of Scedosporiosis and Lomentosporiosis.ACS Omega. 2024 Oct 23;9(44):44815-44824. doi: 10.1021/acsomega.4c08257. eCollection 2024 Nov 5. ACS Omega. 2024. PMID: 39524635 Free PMC article.
-
The Transcription Factors AcuK and AcuM Influence Siderophore Biosynthesis of Aspergillus fumigatus.J Fungi (Basel). 2024 Apr 30;10(5):327. doi: 10.3390/jof10050327. J Fungi (Basel). 2024. PMID: 38786682 Free PMC article.
-
Aspergillus fumigatus biology, immunopathogenicity and drug resistance.Nat Rev Microbiol. 2025 May 2. doi: 10.1038/s41579-025-01180-z. Online ahead of print. Nat Rev Microbiol. 2025. PMID: 40316713 Review.
-
Fungal siderophore metabolism with a focus on Aspergillus fumigatus: impact on biotic interactions and potential translational applications.Essays Biochem. 2023 Sep 13;67(5):829-842. doi: 10.1042/EBC20220252. Essays Biochem. 2023. PMID: 37313590 Free PMC article.
-
The Monothiol Glutaredoxin Grx4 Influences Iron Homeostasis and Virulence in Ustilago maydis.J Fungi (Basel). 2023 Nov 17;9(11):1112. doi: 10.3390/jof9111112. J Fungi (Basel). 2023. PMID: 37998917 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
