

The motor domain of the kinesin Kip2 promotes microtubule polymerization at microtubule tips
- PMID: 37093124
- PMCID: PMC10130750
- DOI: 10.1083/jcb.202110126
The motor domain of the kinesin Kip2 promotes microtubule polymerization at microtubule tips
Abstract
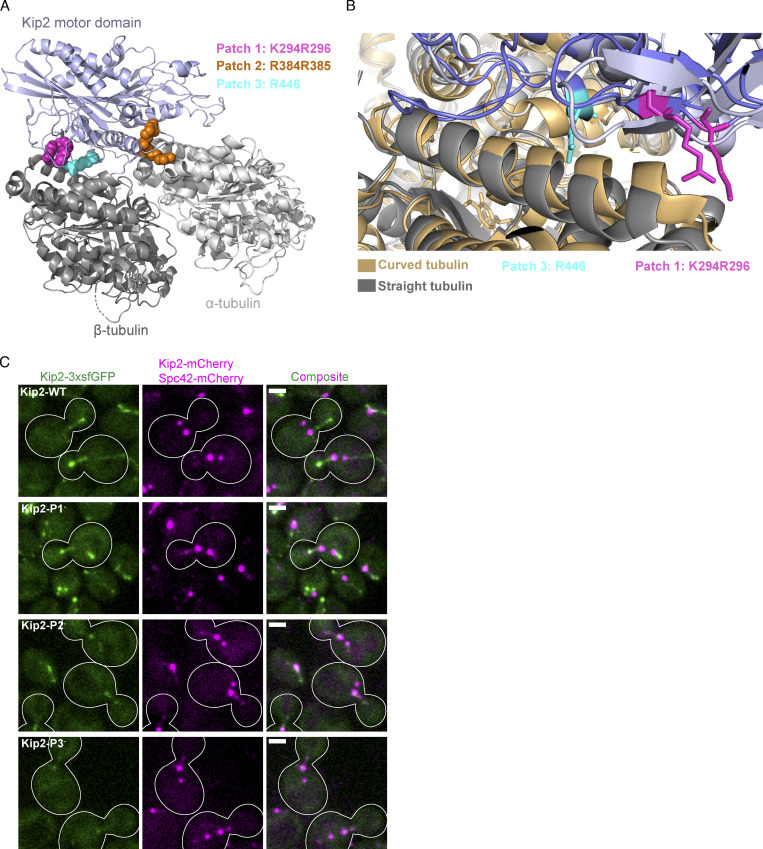
Kinesins are microtubule-dependent motor proteins, some of which moonlight as microtubule polymerases, such as the yeast protein Kip2. Here, we show that the CLIP-170 ortholog Bik1 stabilizes Kip2 at microtubule ends where the motor domain of Kip2 promotes microtubule polymerization. Live-cell imaging and mathematical estimation of Kip2 dynamics reveal that disrupting the Kip2-Bik1 interaction aborts Kip2 dwelling at microtubule ends and abrogates its microtubule polymerization activity. Structural modeling and biochemical experiments identify a patch of positively charged residues that enables the motor domain to bind free tubulin dimers alternatively to the microtubule shaft. Neutralizing this patch abolished the ability of Kip2 to promote microtubule growth both in vivo and in vitro without affecting its ability to walk along microtubules. Our studies suggest that Kip2 utilizes Bik1 as a cofactor to track microtubule tips, where its motor domain then recruits free tubulin and catalyzes microtubule assembly.
© 2023 Chen et al.
Conflict of interest statement
Disclosures: L.A. Widmer reported personal fees from Novartis Pharma AG outside the submitted work. No other disclosures were reported.
Figures











References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
