

Tau in cerebrospinal fluid induces neuronal hyperexcitability and alters hippocampal theta oscillations
- PMID: 37095572
- PMCID: PMC10127378
- DOI: 10.1186/s40478-023-01562-5
Tau in cerebrospinal fluid induces neuronal hyperexcitability and alters hippocampal theta oscillations
Abstract
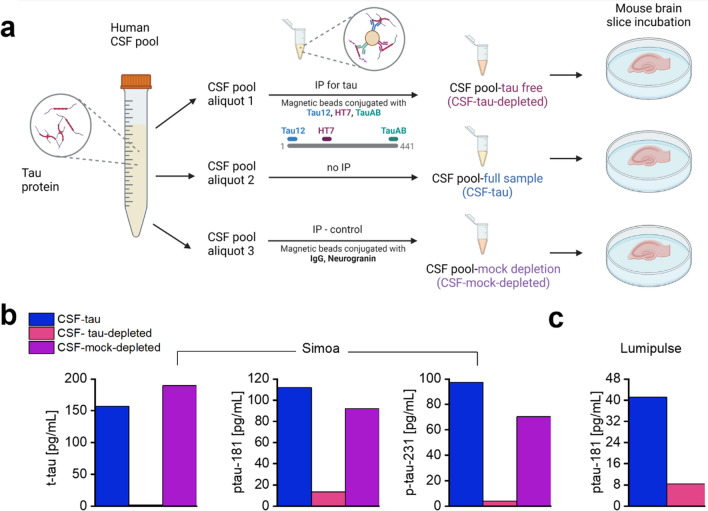
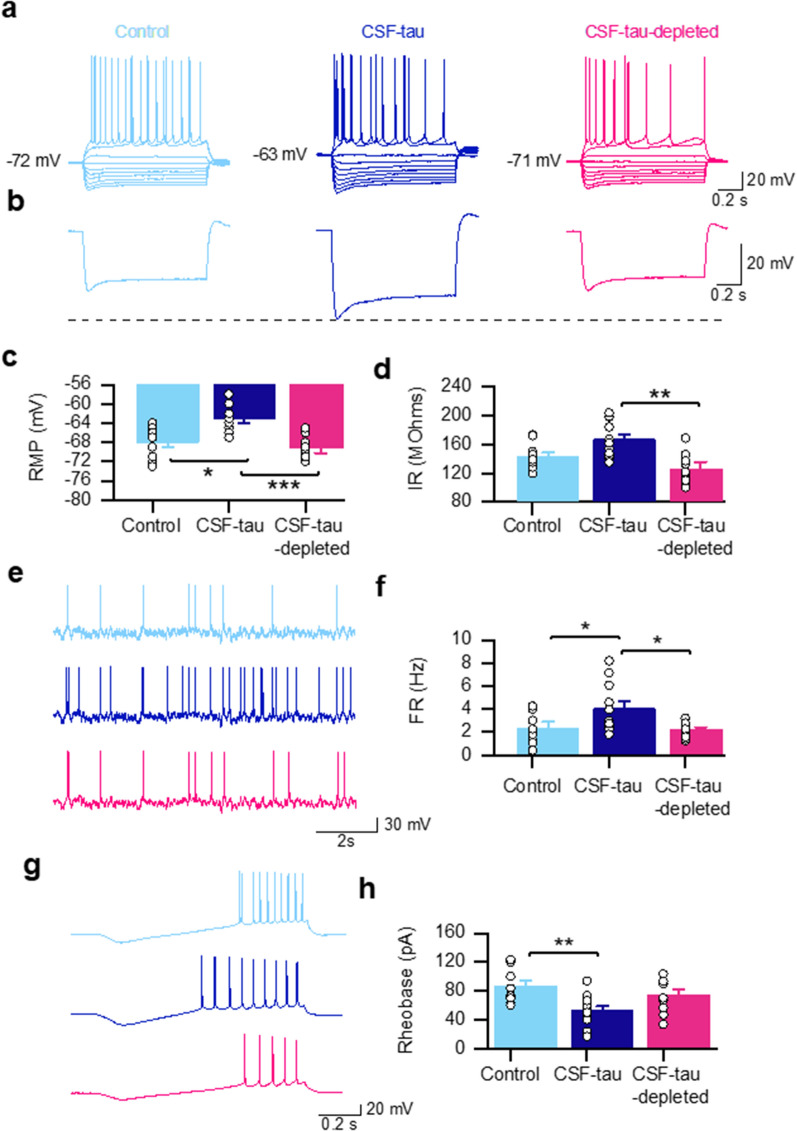
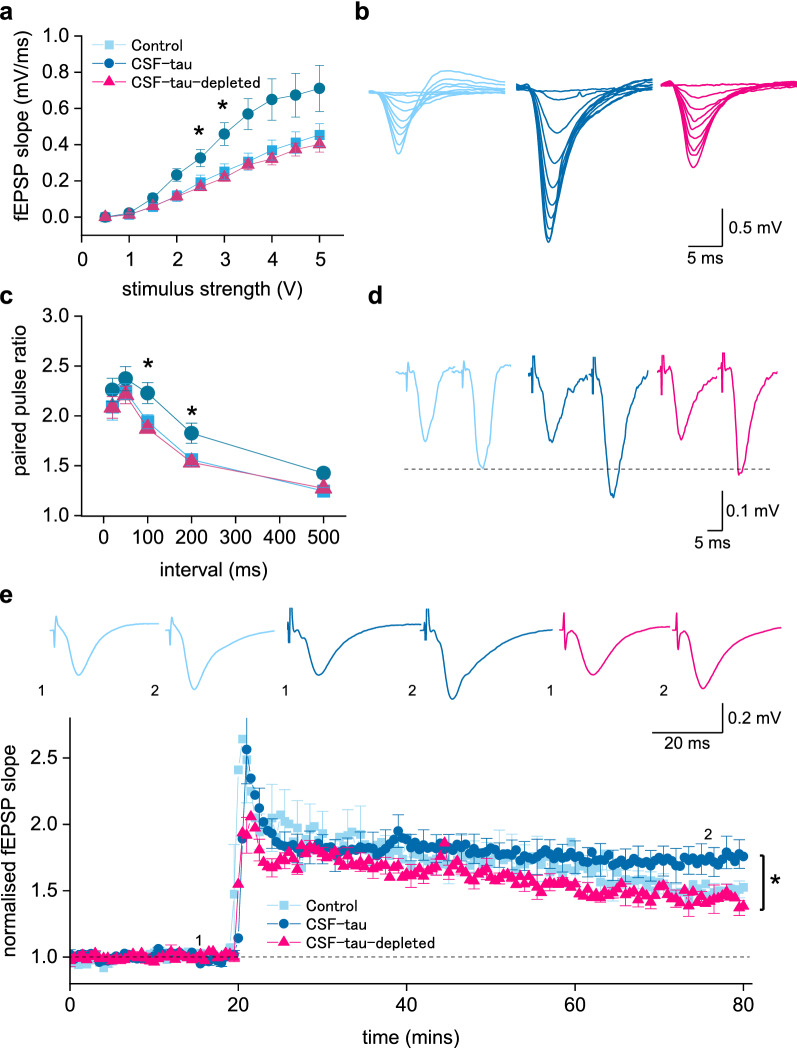
Alzheimer's disease (AD) and other tauopathies are characterized by the aggregation of tau into soluble and insoluble forms (including tangles and neuropil threads). In humans, a fraction of both phosphorylated and non-phosphorylated N-terminal to mid-domain tau species, are secreted into cerebrospinal fluid (CSF). Some of these CSF tau species can be measured as diagnostic and prognostic biomarkers, starting from early stages of disease. While in animal models of AD pathology, soluble tau aggregates have been shown to disrupt neuronal function, it is unclear whether the tau species present in CSF will modulate neural activity. Here, we have developed and applied a novel approach to examine the electrophysiological effects of CSF from patients with a tau-positive biomarker profile. The method involves incubation of acutely-isolated wild-type mouse hippocampal brain slices with small volumes of diluted human CSF, followed by a suite of electrophysiological recording methods to evaluate their effects on neuronal function, from single cells through to the network level. Comparison of the toxicity profiles of the same CSF samples, with and without immuno-depletion for tau, has enabled a pioneering demonstration that CSF-tau potently modulates neuronal function. We demonstrate that CSF-tau mediates an increase in neuronal excitability in single cells. We then observed, at the network level, increased input-output responses and enhanced paired-pulse facilitation as well as an increase in long-term potentiation. Finally, we show that CSF-tau modifies the generation and maintenance of hippocampal theta oscillations, which have important roles in learning and memory and are known to be altered in AD patients. Together, we describe a novel method for screening human CSF-tau to understand functional effects on neuron and network activity, which could have far-reaching benefits in understanding tau pathology, thus allowing for the development of better targeted treatments for tauopathies in the future.
Keywords: Cerebrospinal fluid; Electrophysiology; Tau; Tauopathy; Theta oscillations.
© 2023. The Author(s).
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures


Similar articles
-
Phospho-tau serine-262 and serine-356 as biomarkers of pre-tangle soluble tau assemblies in Alzheimer's disease.Nat Med. 2025 Feb;31(2):574-588. doi: 10.1038/s41591-024-03400-0. Epub 2025 Feb 10. Nat Med. 2025. PMID: 39930142 Free PMC article.
-
Cerebrospinal fluid tau fragment correlates with tau PET: a candidate biomarker for tangle pathology.Brain. 2020 Feb 1;143(2):650-660. doi: 10.1093/brain/awz346. Brain. 2020. PMID: 31834365 Free PMC article.
-
Divergent CSF τ alterations in two common tauopathies: Alzheimer's disease and progressive supranuclear palsy.J Neurol Neurosurg Psychiatry. 2015 Mar;86(3):244-50. doi: 10.1136/jnnp-2014-308004. Epub 2014 Jun 4. J Neurol Neurosurg Psychiatry. 2015. PMID: 24899730 Free PMC article.
-
Cerebrospinal Fluid and Plasma Tau as a Biomarker for Brain Tauopathy.Adv Exp Med Biol. 2019;1184:393-405. doi: 10.1007/978-981-32-9358-8_29. Adv Exp Med Biol. 2019. PMID: 32096052 Review.
-
Clinical utility and analytical challenges in measurement of cerebrospinal fluid amyloid-β(1-42) and τ proteins as Alzheimer disease biomarkers.Clin Chem. 2013 Jun;59(6):903-16. doi: 10.1373/clinchem.2013.202937. Epub 2013 Mar 21. Clin Chem. 2013. PMID: 23519967 Free PMC article. Review.
Cited by
-
Anti-epileptic drug use and subsequent degenerative dementia occurrence.Alzheimers Dement (N Y). 2024 Sep 10;10(3):e70001. doi: 10.1002/trc2.70001. eCollection 2024 Jul-Sep. Alzheimers Dement (N Y). 2024. PMID: 39257557 Free PMC article.
-
Pathological tau alters head direction signaling and induces spatial disorientation.bioRxiv [Preprint]. 2025 Feb 25:2024.11.07.622548. doi: 10.1101/2024.11.07.622548. bioRxiv. 2025. PMID: 39574637 Free PMC article. Preprint.
-
Loss of excitatory inputs and decreased tonic and evoked activity of locus coeruleus neurons in aged P301S mice.bioRxiv [Preprint]. 2025 Jan 17:2025.01.17.633373. doi: 10.1101/2025.01.17.633373. bioRxiv. 2025. Update in: Neurobiol Dis. 2025 May;208:106883. doi: 10.1016/j.nbd.2025.106883. PMID: 39868303 Free PMC article. Updated. Preprint.
-
Rapidly reversible persistent long-term potentiation inhibition by patient-derived brain tau and amyloid ß proteins.Philos Trans R Soc Lond B Biol Sci. 2024 Jul 29;379(1906):20230234. doi: 10.1098/rstb.2023.0234. Epub 2024 Jun 10. Philos Trans R Soc Lond B Biol Sci. 2024. PMID: 38853565 Free PMC article.
-
Immunoassay for pyruvate kinase M1/2 as an Alzheimer's biomarker in CSF.Open Life Sci. 2025 Jul 7;20(1):20251101. doi: 10.1515/biol-2025-1101. eCollection 2025. Open Life Sci. 2025. PMID: 40667488 Free PMC article.
References
-
- Acquarone E, Argyrousi EK, van den Berg M, Gulisano W, Fà M, Staniszewski A, Calcagno E, Zuccarello E, D’Adamio L, Deng S-X, Puzzo D, Arancio O, Fiorito J. Synaptic and memory dysfunction induced by tau oligomers is rescued by up-regulation of the nitric oxide cascade. Mol Neurodegener. 2019;14:26. doi: 10.1186/s13024-019-0326-4. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
