

GIPC3 couples to MYO6 and PDZ domain proteins, and shapes the hair cell apical region
- PMID: 37096733
- PMCID: PMC10234109
- DOI: 10.1242/jcs.261100
GIPC3 couples to MYO6 and PDZ domain proteins, and shapes the hair cell apical region
Abstract
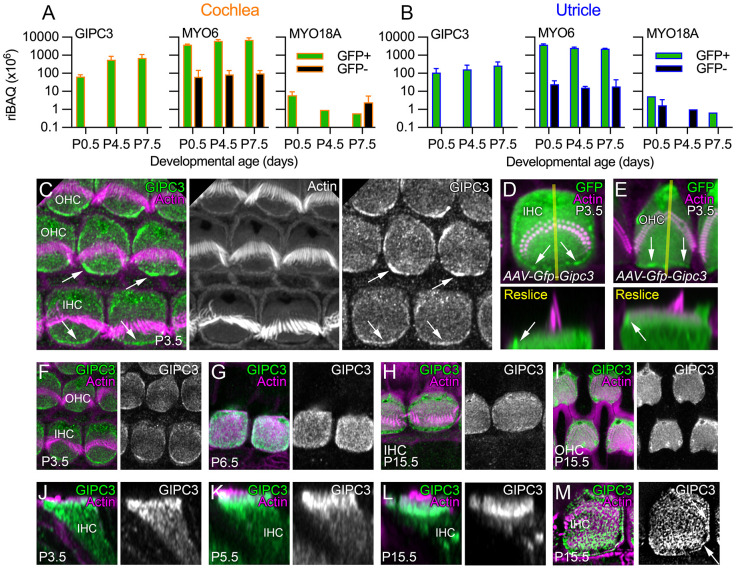
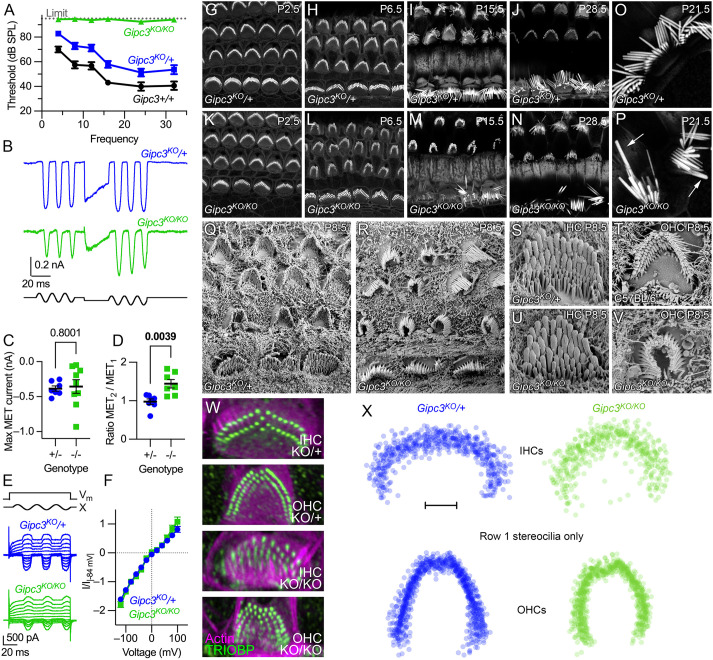
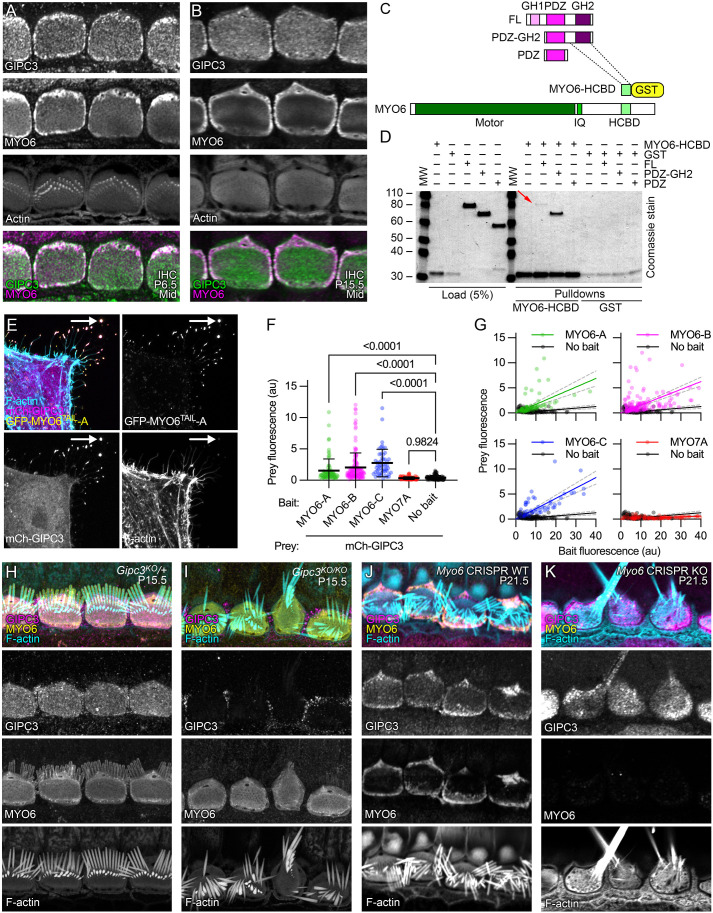
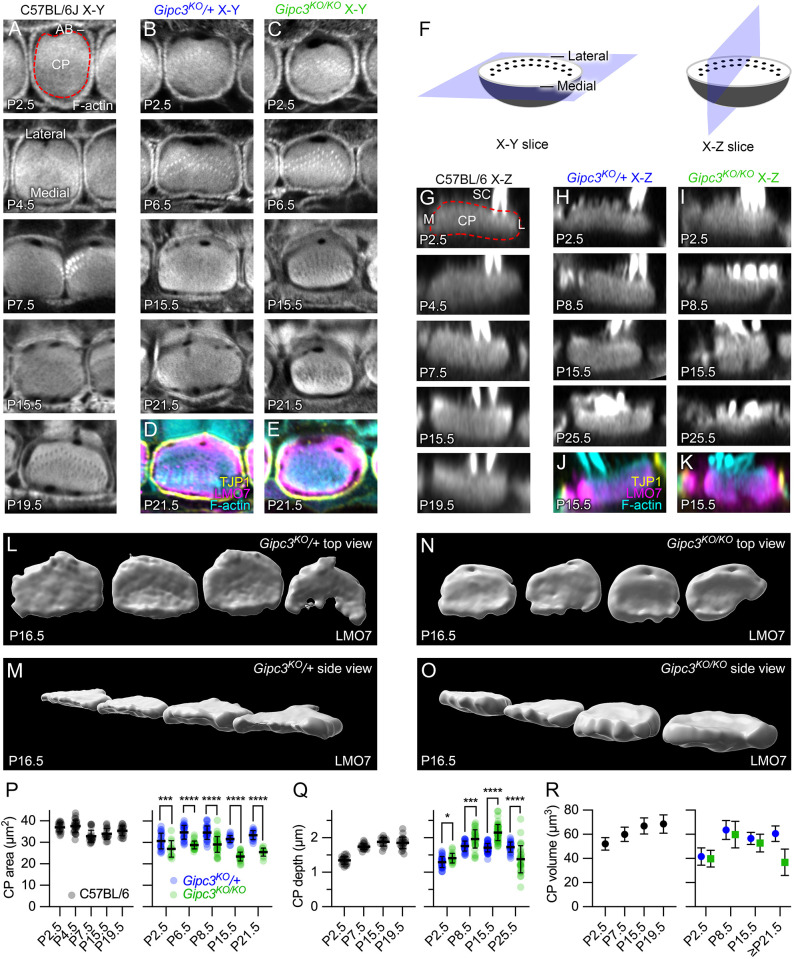
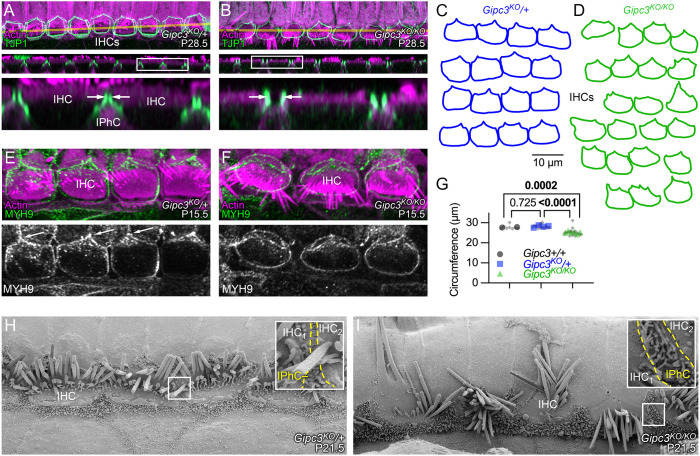
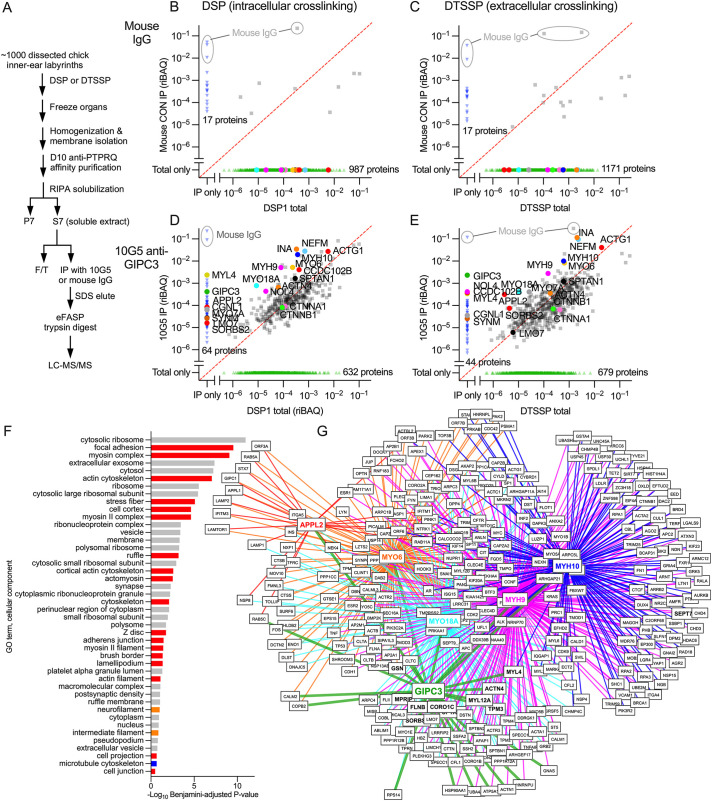
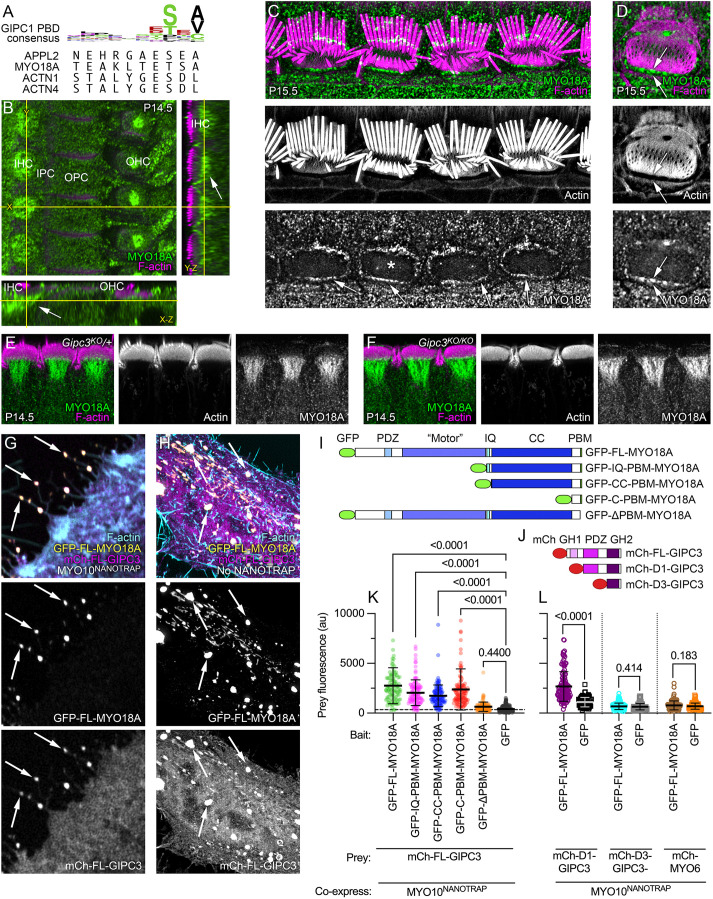
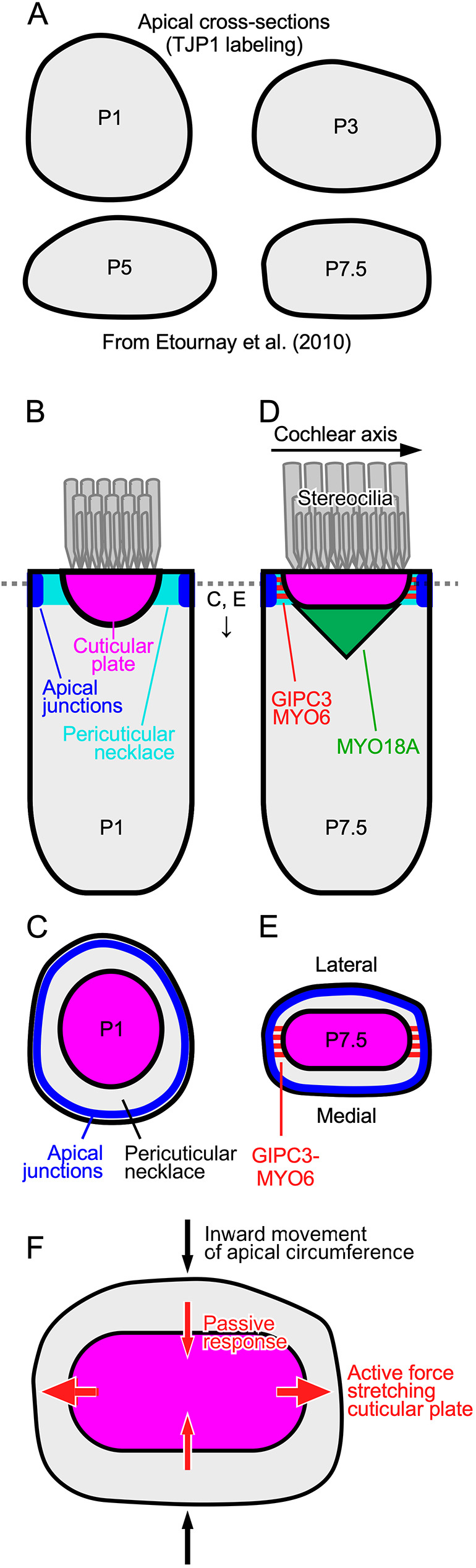
GIPC3 has been implicated in auditory function. Here, we establish that GIPC3 is initially localized to the cytoplasm of inner and outer hair cells of the cochlea and then is increasingly concentrated in cuticular plates and at cell junctions during postnatal development. Early postnatal Gipc3KO/KO mice had mostly normal mechanotransduction currents, but had no auditory brainstem response at 1 month of age. Cuticular plates of Gipc3KO/KO hair cells did not flatten during development as did those of controls; moreover, hair bundles were squeezed along the cochlear axis in mutant hair cells. Junctions between inner hair cells and adjacent inner phalangeal cells were also severely disrupted in Gipc3KO/KO cochleas. GIPC3 bound directly to MYO6, and the loss of MYO6 led to altered distribution of GIPC3. Immunoaffinity purification of GIPC3 from chicken inner ear extracts identified co-precipitating proteins associated with adherens junctions, intermediate filament networks and the cuticular plate. Several of immunoprecipitated proteins contained GIPC family consensus PDZ-binding motifs (PBMs), including MYO18A, which bound directly to the PDZ domain of GIPC3. We propose that GIPC3 and MYO6 couple to PBMs of cytoskeletal and cell junction proteins to shape the cuticular plate.
Keywords: Actin; Cuticular plate; Cytoskeleton; Hair cell; Myosin.
© 2023. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
Update of
-
GIPC3 couples to MYO6 and PDZ domain proteins and shapes the hair cell apical region.bioRxiv [Preprint]. 2023 Mar 1:2023.02.28.530466. doi: 10.1101/2023.02.28.530466. bioRxiv. 2023. Update in: J Cell Sci. 2023 May 15;136(10):jcs261100. doi: 10.1242/jcs.261100. PMID: 36909580 Free PMC article. Updated. Preprint.
Similar articles
-
GIPC3 couples to MYO6 and PDZ domain proteins and shapes the hair cell apical region.bioRxiv [Preprint]. 2023 Mar 1:2023.02.28.530466. doi: 10.1101/2023.02.28.530466. bioRxiv. 2023. Update in: J Cell Sci. 2023 May 15;136(10):jcs261100. doi: 10.1242/jcs.261100. PMID: 36909580 Free PMC article. Updated. Preprint.
-
A novel splice site mutation of myosin VI in mice leads to stereociliary fusion caused by disruption of actin networks in the apical region of inner ear hair cells.PLoS One. 2017 Aug 23;12(8):e0183477. doi: 10.1371/journal.pone.0183477. eCollection 2017. PLoS One. 2017. PMID: 28832620 Free PMC article.
-
Gipc3 mutations associated with audiogenic seizures and sensorineural hearing loss in mouse and human.Nat Commun. 2011 Feb 15;2:201. doi: 10.1038/ncomms1200. Nat Commun. 2011. PMID: 21326233 Free PMC article.
-
The cuticular plate: a riddle, wrapped in a mystery, inside a hair cell.Birth Defects Res C Embryo Today. 2015 Jun;105(2):126-39. doi: 10.1002/bdrc.21098. Epub 2015 Jun 23. Birth Defects Res C Embryo Today. 2015. PMID: 26104653 Review.
-
Functional proteomics, human genetics and cancer biology of GIPC family members.Exp Mol Med. 2013 Jun 7;45(6):e26. doi: 10.1038/emm.2013.49. Exp Mol Med. 2013. PMID: 23743496 Free PMC article. Review.
Cited by
-
The genomic landscape of Ménière's disease: a path to endolymphatic hydrops.BMC Genomics. 2024 Jun 28;25(1):646. doi: 10.1186/s12864-024-10552-3. BMC Genomics. 2024. PMID: 38943082 Free PMC article.
-
Are the class 18 myosins Myo18A and Myo18B specialist sarcomeric proteins?Front Physiol. 2024 May 9;15:1401717. doi: 10.3389/fphys.2024.1401717. eCollection 2024. Front Physiol. 2024. PMID: 38784114 Free PMC article. Review.
-
Single-molecule fluorescence microscopy reveals regulatory mechanisms of MYO7A-driven cargo transport in stereocilia of live inner ear hair cells.Nat Commun. 2025 Sep 1;16(1):8149. doi: 10.1038/s41467-025-63102-0. Nat Commun. 2025. PMID: 40890108 Free PMC article.
-
Microanatomy of the human tunnel of Corti structures and cochlear partition-tonotopic variations and transcellular signaling.J Anat. 2024 Aug;245(2):271-288. doi: 10.1111/joa.14045. Epub 2024 Apr 13. J Anat. 2024. PMID: 38613211 Free PMC article.
-
Pathophysiology of human hearing loss associated with variants in myosins.Front Physiol. 2024 Mar 18;15:1374901. doi: 10.3389/fphys.2024.1374901. eCollection 2024. Front Physiol. 2024. PMID: 38562617 Free PMC article. Review.
References
-
- Avraham, K. B., Hasson, T., Sobe, T., Balsara, B., Testa, J. R., Skvorak, A. B., Morton, C. C., Copeland, N. G. and Jenkins, N. A. (1997). Characterization of unconventional MYO6, the human homologue of the gene responsible for deafness in Snell's waltzer mice. Hum. Mol. Genet. 6, 1225-1231. 10.1093/hmg/6.8.1225 - DOI - PubMed
-
- Bird, J. E., Barzik, M., Drummond, M. C., Sutton, D. C., Goodman, S. M., Morozko, E. L., Cole, S. M., Boukhvalova, A. K., Skidmore, J., Syam, D., et al. (2017). Harnessing molecular motors for nanoscale pulldown in live cells. Mol. Biol. Cell 28, 463-475. 10.1091/mbc.e16-08-0583 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
