

The Timing and Magnitude of the Type I Interferon Response Are Correlated with Disease Tolerance in Arbovirus Infection
- PMID: 37097030
- PMCID: PMC10294695
- DOI: 10.1128/mbio.00101-23
The Timing and Magnitude of the Type I Interferon Response Are Correlated with Disease Tolerance in Arbovirus Infection
Abstract
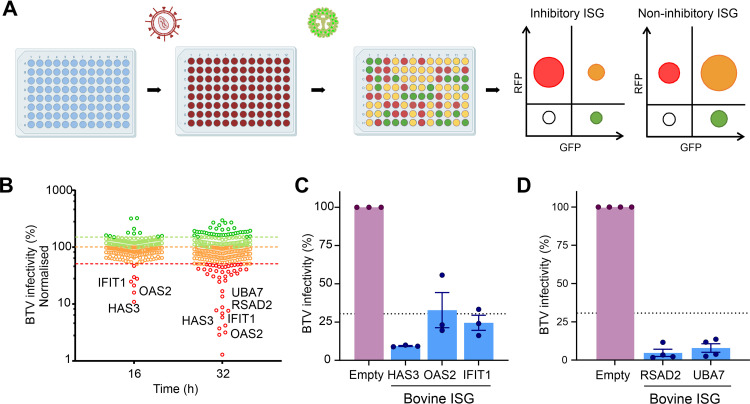
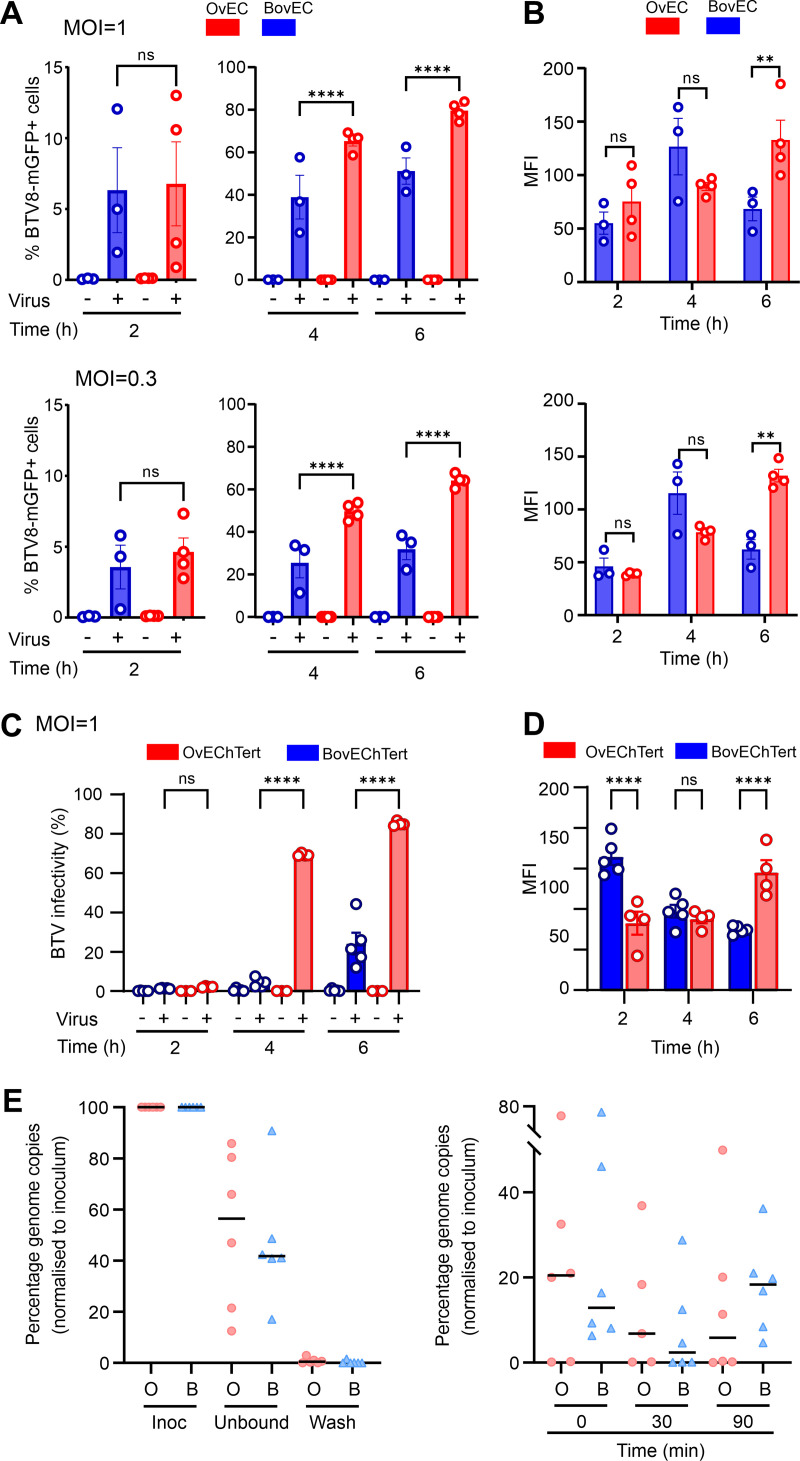
Infected hosts possess two alternative strategies to protect themselves against the negative impact of virus infections: resistance, used to abrogate virus replication, and disease tolerance, used to avoid tissue damage without controlling viral burden. The principles governing pathogen resistance are well understood, while less is known about those involved in disease tolerance. Here, we studied bluetongue virus (BTV), the cause of bluetongue disease of ruminants, as a model system to investigate the mechanisms of virus-host interactions correlating with disease tolerance. BTV induces clinical disease mainly in sheep, while cattle are considered reservoirs of infection, rarely exhibiting clinical symptoms despite sustained viremia. Using primary cells from multiple donors, we show that BTV consistently reaches higher titers in ovine cells than cells from cattle. The variable replication kinetics of BTV in sheep and cow cells were mostly abolished by abrogating the cell type I interferon (IFN) response. We identified restriction factors blocking BTV replication, but both the sheep and cow orthologues of these antiviral genes possess anti-BTV properties. Importantly, we demonstrate that BTV induces a faster host cell protein synthesis shutoff in primary sheep cells than cow cells, which results in an earlier downregulation of antiviral proteins. Moreover, by using RNA sequencing (RNA-seq), we also show a more pronounced expression of interferon-stimulated genes (ISGs) in BTV-infected cow cells than sheep cells. Our data provide a new perspective on how the type I IFN response in reservoir species can have overall positive effects on both virus and host evolution. IMPORTANCE The host immune response usually aims to inhibit virus replication in order to avoid cell damage and disease. In some cases, however, the infected host avoids the deleterious effects of infection despite high levels of viral replication. This strategy is known as disease tolerance, and it is used by animal reservoirs of some zoonotic viruses. Here, using a virus of ruminants (bluetongue virus [BTV]) as an experimental system, we dissected virus-host interactions in cells collected from species that are susceptible (sheep) or tolerant (cow) to disease. We show that (i) virus modulation of the host antiviral type I interferon (IFN) responses, (ii) viral replication kinetics, and (iii) virus-induced cell damage differ in tolerant and susceptible BTV-infected cells. Understanding the complex virus-host interactions in disease tolerance can allow us to disentangle the critical balance between protective and damaging host immune responses.
Keywords: arbovirus; disease tolerance; innate immunity; interferons.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
Bluetongue Virus NS4 Protein Is an Interferon Antagonist and a Determinant of Virus Virulence.J Virol. 2016 May 12;90(11):5427-39. doi: 10.1128/JVI.00422-16. Print 2016 Jun 1. J Virol. 2016. PMID: 27009961 Free PMC article.
-
Dual modulation of type I interferon response by bluetongue virus.J Virol. 2014 Sep;88(18):10792-802. doi: 10.1128/JVI.01235-14. Epub 2014 Jul 9. J Virol. 2014. PMID: 25008919 Free PMC article.
-
Comparative effects of bluetongue virus infection of ovine and bovine endothelial cells.Vet Pathol. 1996 May;33(3):319-31. doi: 10.1177/030098589603300309. Vet Pathol. 1996. PMID: 8740706
-
[Advances in the anti-host interferon immune response of bluetongue virus].Sheng Wu Gong Cheng Xue Bao. 2024 Dec 25;40(12):4439-4451. doi: 10.13345/j.cjb.240080. Sheng Wu Gong Cheng Xue Bao. 2024. PMID: 39722507 Review. Chinese.
-
Vaccines for Prevention of Bluetongue and Epizootic Hemorrhagic Disease in Livestock: A North American Perspective.Vector Borne Zoonotic Dis. 2015 Jun;15(6):385-96. doi: 10.1089/vbz.2014.1698. Vector Borne Zoonotic Dis. 2015. PMID: 26086559 Review.
Cited by
-
Comprehensive immune profiling reveals that Orbivirus infection activates immune checkpoints during acute T cell immunosuppression.Front Immunol. 2023 Oct 18;14:1255803. doi: 10.3389/fimmu.2023.1255803. eCollection 2023. Front Immunol. 2023. PMID: 37920474 Free PMC article.
-
Engineering recombinant replication-competent bluetongue viruses expressing reporter genes for in vitro and non-invasive in vivo studies.Microbiol Spectr. 2024 Mar 5;12(3):e0249323. doi: 10.1128/spectrum.02493-23. Epub 2024 Feb 14. Microbiol Spectr. 2024. PMID: 38353566 Free PMC article.
-
Bluetongue's New Frontier-Are Dogs at Risk?Vet Sci. 2025 May 20;12(5):505. doi: 10.3390/vetsci12050505. Vet Sci. 2025. PMID: 40431598 Free PMC article. Review.
-
Defining correlates of protection for mammalian livestock vaccines against high-priority viral diseases.Front Immunol. 2024 Jul 19;15:1397780. doi: 10.3389/fimmu.2024.1397780. eCollection 2024. Front Immunol. 2024. PMID: 39100679 Free PMC article. Review.
-
Correlates of disease severity in bluetongue as a model of acute arbovirus infection.PLoS Pathog. 2024 Aug 16;20(8):e1012466. doi: 10.1371/journal.ppat.1012466. eCollection 2024 Aug. PLoS Pathog. 2024. PMID: 39150989 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
