

CD30 co-stimulation drives differentiation of protective T cells during Mycobacterium tuberculosis infection
- PMID: 37097292
- PMCID: PMC10130742
- DOI: 10.1084/jem.20222090
CD30 co-stimulation drives differentiation of protective T cells during Mycobacterium tuberculosis infection
Abstract
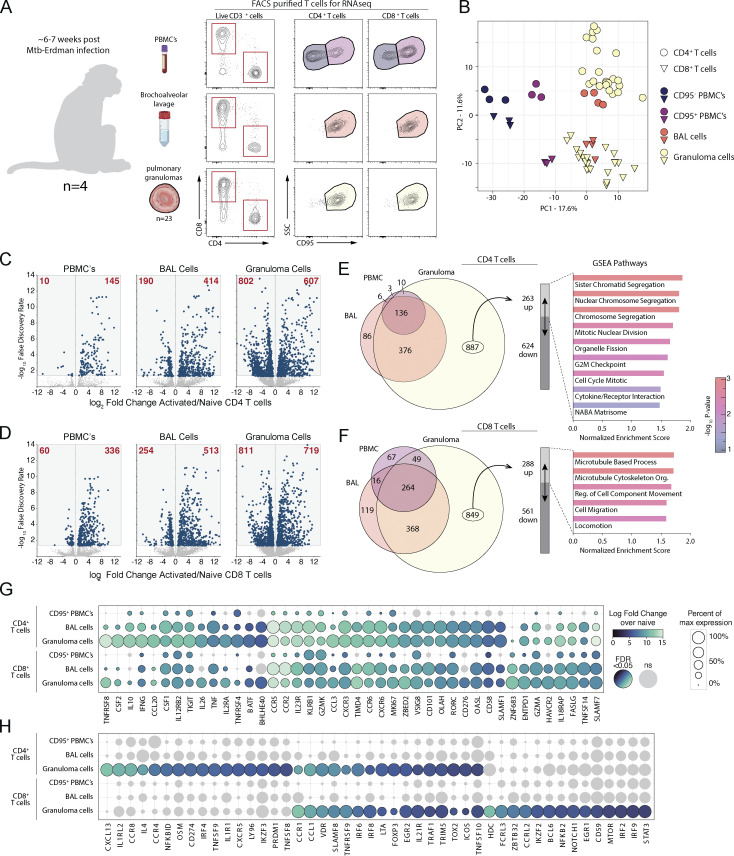
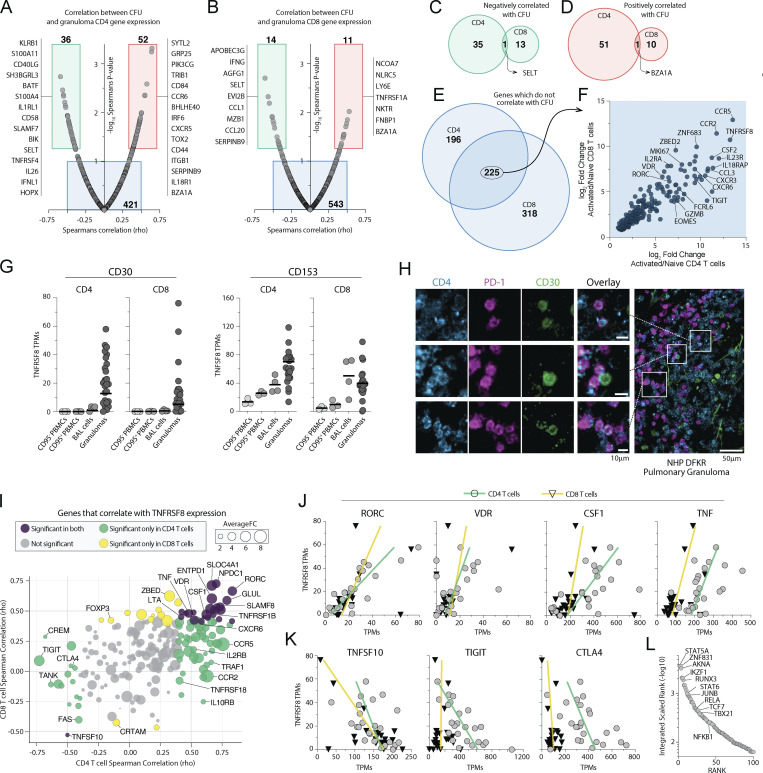
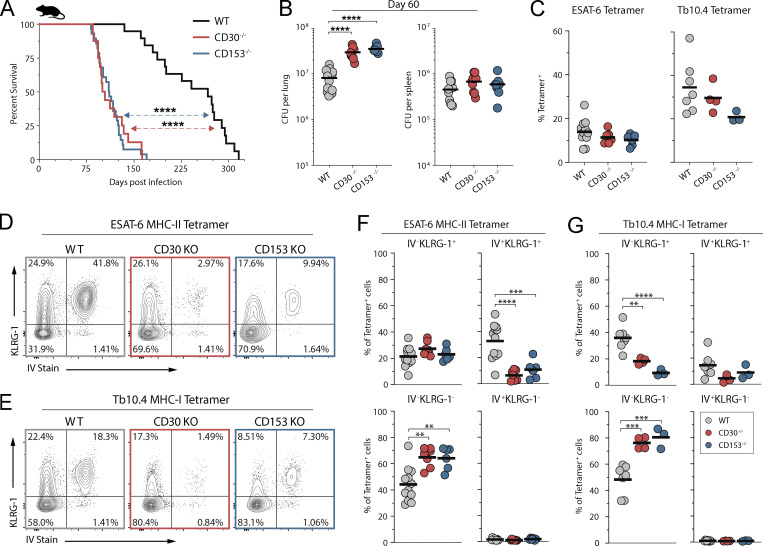
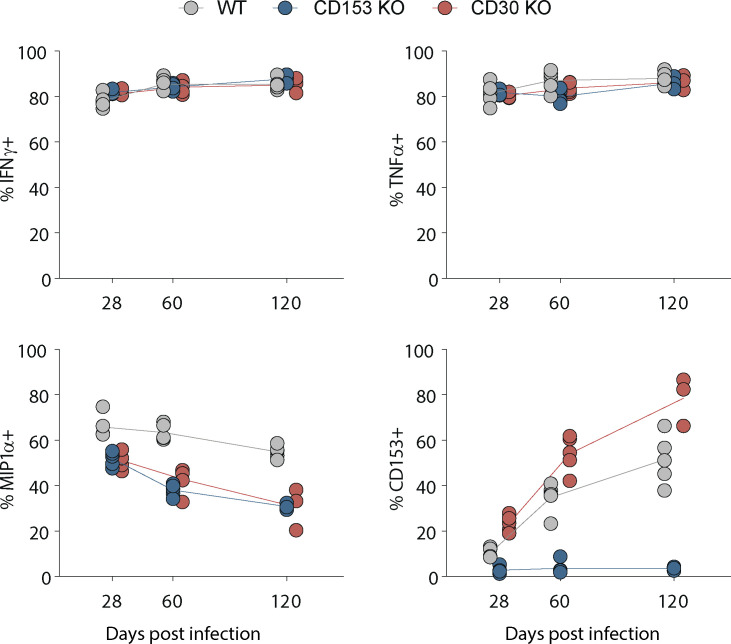
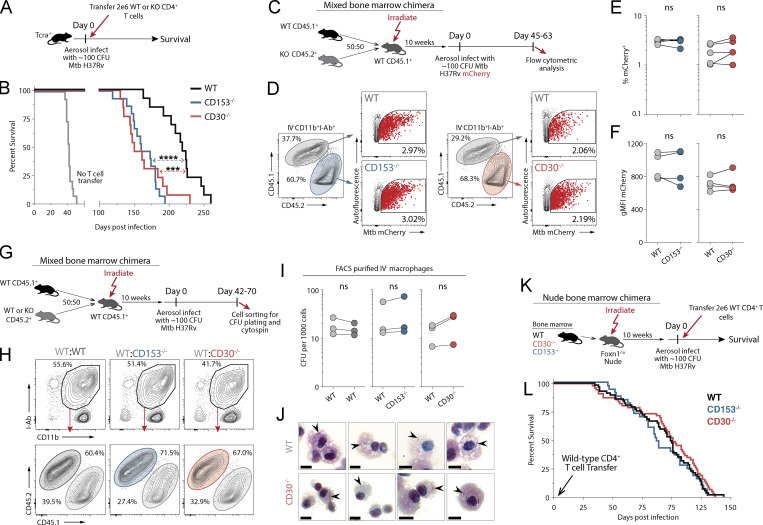
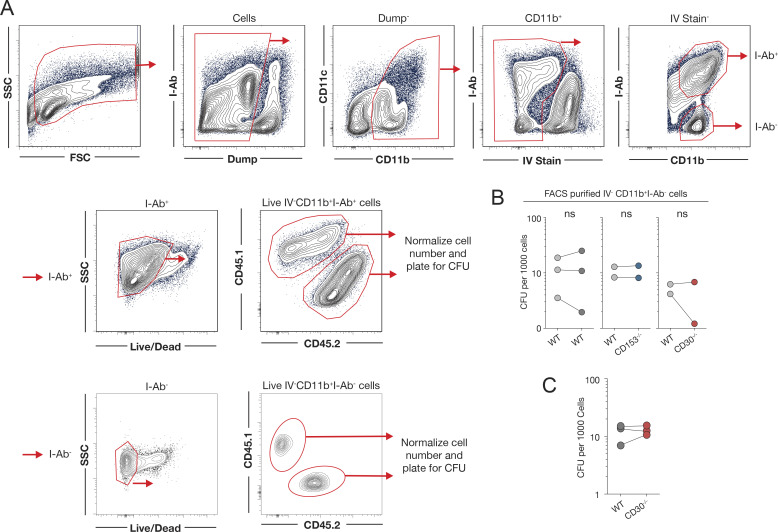
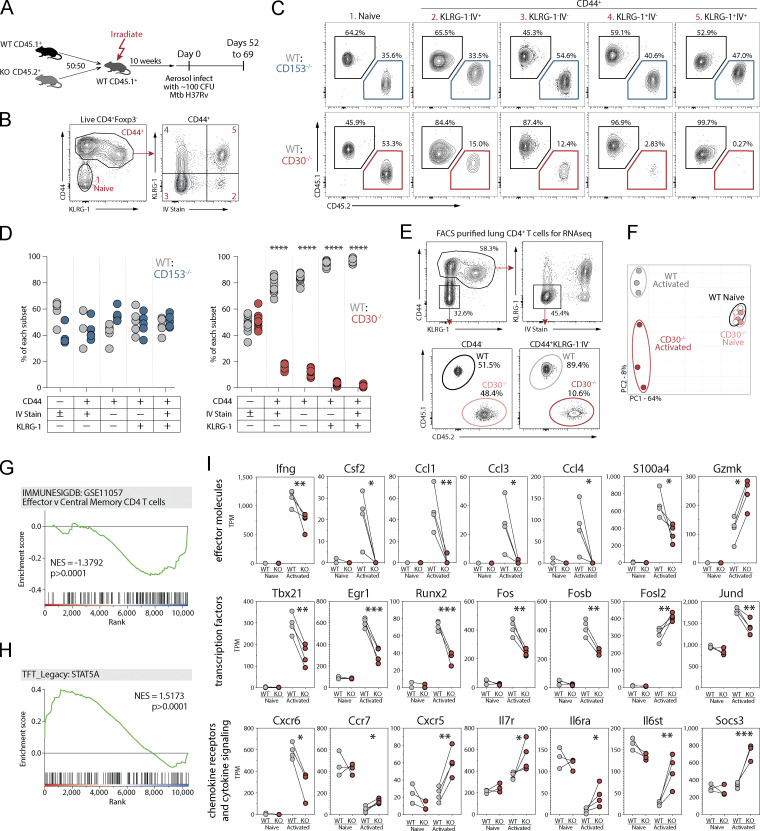
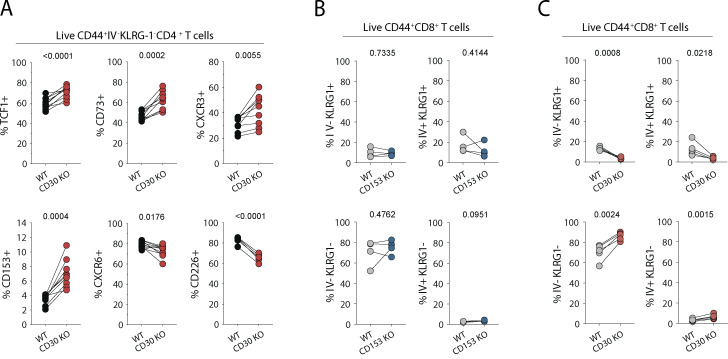
Control of Mycobacterium tuberculosis (Mtb) infection requires generation of T cells that migrate to granulomas, complex immune structures surrounding sites of bacterial replication. Here we compared the gene expression profiles of T cells in pulmonary granulomas, bronchoalveolar lavage, and blood of Mtb-infected rhesus macaques to identify granuloma-enriched T cell genes. TNFRSF8/CD30 was among the top genes upregulated in both CD4 and CD8 T cells from granulomas. In mice, CD30 expression on CD4 T cells is required for survival of Mtb infection, and there is no major role for CD30 in protection by other cell types. Transcriptomic comparison of WT and CD30-/- CD4 T cells from the lungs of Mtb-infected mixed bone marrow chimeric mice showed that CD30 directly promotes CD4 T cell differentiation and the expression of multiple effector molecules. These results demonstrate that the CD30 co-stimulatory axis is highly upregulated on granuloma T cells and is critical for protective T cell responses against Mtb infection.
This is a work of the U.S. Government and is not subject to copyright protection in the United States. Foreign copyrights may apply.
Conflict of interest statement
Disclosures: T.W. Foreman, K.D. Kauffman, M.A. Sallin, and D.L. Barber reported a patent to CD153 and/or CD30 in infection, application no. 62/633,816, pending. No other disclosures were reported.
Figures
Comment in
-
TB granuloma: CD30 co-stimulation for CD4+ T cell co-operation.J Exp Med. 2023 Aug 7;220(8):e20230547. doi: 10.1084/jem.20230547. Epub 2023 May 9. J Exp Med. 2023. PMID: 37158981 Free PMC article.
References
-
- Burel, J.G., Lindestam Arlehamn C.S., Khan N., Seumois G., Greenbaum J.A., Taplitz R., Gilman R.H., Saito M., Vijayanand P., Sette A., and Peters B.. 2018. Transcriptomic Analysis of CD4(+) T Cells Reveals Novel Immune Signatures of Latent Tuberculosis. J. Immunol. 200:3283–3290. 10.4049/jimmunol.1800118 - DOI - PMC - PubMed
-
- Chen, Y., Zander R.A., Wu X., Schauder D.M., Kasmani M.Y., Shen J., Zheng S., Burns R., Taparowsky E.J., and Cui W.. 2021. BATF regulates progenitor to cytolytic effector CD8+ T cell transition during chronic viral infection. Nat. Immunol. 22:996–1007. 10.1038/s41590-021-00965-7 - DOI - PMC - PubMed
-
- Clemmensen, H.S., Dube J.Y., McIntosh F., Rosenkrands I., Jungersen G., Aagaard C., Andersen P., Behr M.A., and Mortensen R.. 2021. In Vivo Antigen Expression Regulates CD4 T Cell Differentiation and Vaccine Efficacy against Mycobacterium tuberculosis Infection. mBio. 12. e00226-21. 10.1128/mBio.00226-21 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
