

Geometry-mediated bridging drives nonadhesive stripe wound healing
- PMID: 37098071
- PMCID: PMC10161107
- DOI: 10.1073/pnas.2221040120
Geometry-mediated bridging drives nonadhesive stripe wound healing
Abstract
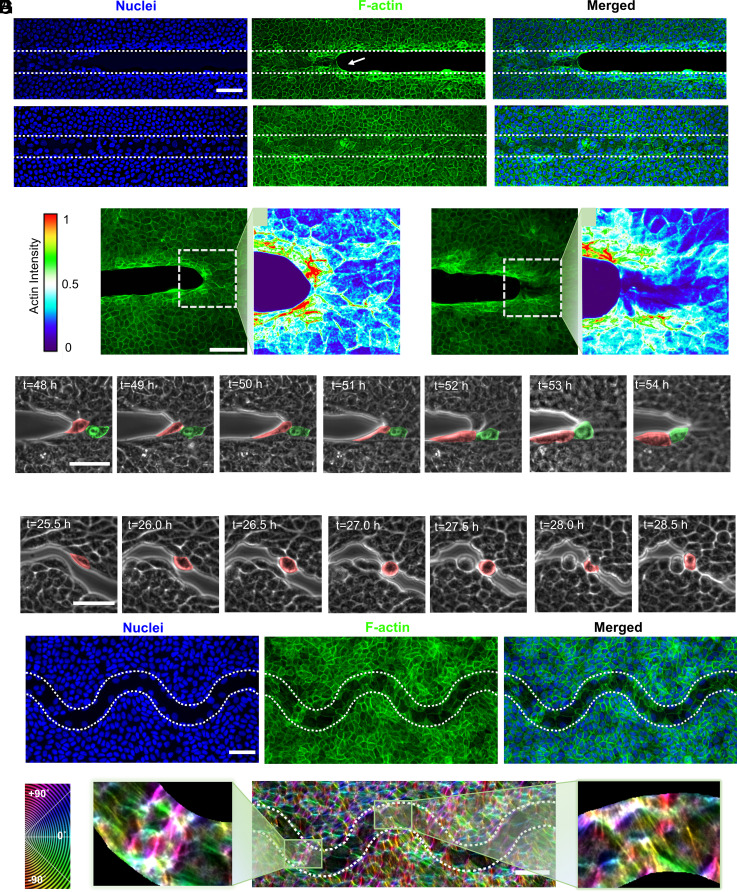
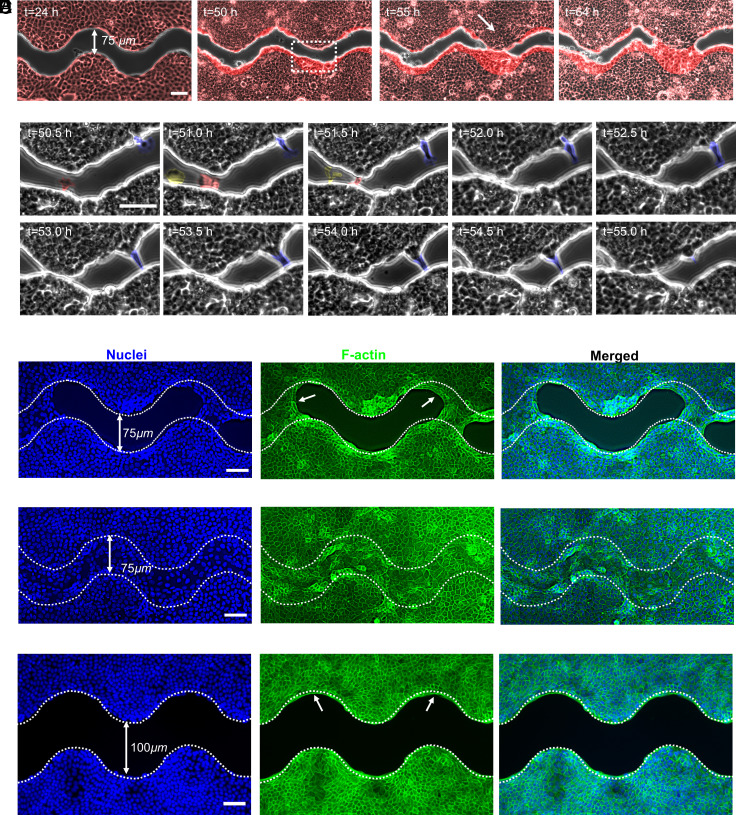
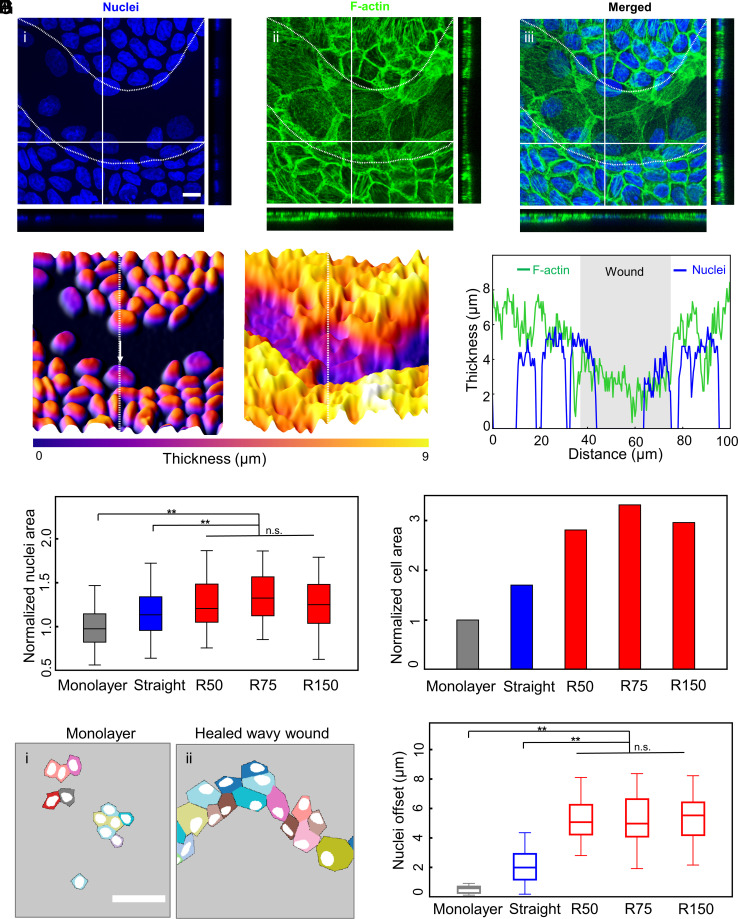
Wound healing through reepithelialization of gaps is of profound importance to the medical community. One critical mechanism identified by researchers for closing non-cell-adhesive gaps is the accumulation of actin cables around concave edges and the resulting purse-string constriction. However, the studies to date have not separated the gap-edge curvature effect from the gap size effect. Here, we fabricate micropatterned hydrogel substrates with long, straight, and wavy non-cell-adhesive stripes of different gap widths to investigate the stripe edge curvature and stripe width effects on the reepithelialization of Madin-Darby canine kidney (MDCK) cells. Our results show that MDCK cell reepithelization is closely regulated by the gap geometry and may occur through different pathways. In addition to purse-string contraction, we identify gap bridging either via cell protrusion or by lamellipodium extension as critical cellular and molecular mechanisms for wavy gap closure. Cell migration in the direction perpendicular to wound front, sufficiently small gap size to allow bridging, and sufficiently high negative curvature at cell bridges for actin cable constriction are necessary/sufficient conditions for gap closure. Our experiments demonstrate that straight stripes rarely induce cell migration perpendicular to wound front, but wavy stripes do; cell protrusion and lamellipodia extension can help establish bridges over gaps of about five times the cell size, but not significantly beyond. Such discoveries deepen our understanding of mechanobiology of cell responses to curvature and help guide development of biophysical strategies for tissue repair, plastic surgery, and better wound management.
Keywords: actin cable; cell protrusion; collective cell migration; gap closure; lamellipodia extension.
Conflict of interest statement
The authors declare no competing interest.
Figures



References
-
- Langer K., On the anatomy and physiology of the skin: I. The cleavability of the cutis. Br. J. Plast. Surg. 31, 3–8 (1978). - PubMed
-
- Azmat C. E., Council M., Wound Closure Techniques (StatPearls Publishing, 2022) (July 17, 2022). - PubMed
-
- Saiko G., “The impact of the wound shape on wound healing dynamics: Is it time to revisit wound healing measures?” in Proceedings of the 14th International Joint Conference on Biomedical Engineering Systems and Technologies, Lorenz R., Fred A., Gamboa H., Eds. (SCITEPRESS - Science and Technology Publications, 2021), pp. 182–187.
-
- Borges A. F., Zigzag incisions for improved exposure and scarring. Clin. Orthop. 145, 202–207 (1979). - PubMed
-
- Nguyen D. T., Orgill D. P., Murphy G. F., “The pathophysiologic basis for wound healing and cutaneous regeneration” in Biomaterials for Treating Skin Loss, Orgill D., Blanco C., Eds. (Woodhead Publishing, Woodhead Publishing Series in Biomaterials, 2009), pp. 25–57.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
