

Never in mitosis gene A-related kinase-8 promotes proliferation, migration, invasion, and stemness of breast cancer cells via β-catenin signalling activation
- PMID: 37100815
- PMCID: PMC10133229
- DOI: 10.1038/s41598-023-32631-3
Never in mitosis gene A-related kinase-8 promotes proliferation, migration, invasion, and stemness of breast cancer cells via β-catenin signalling activation
Abstract
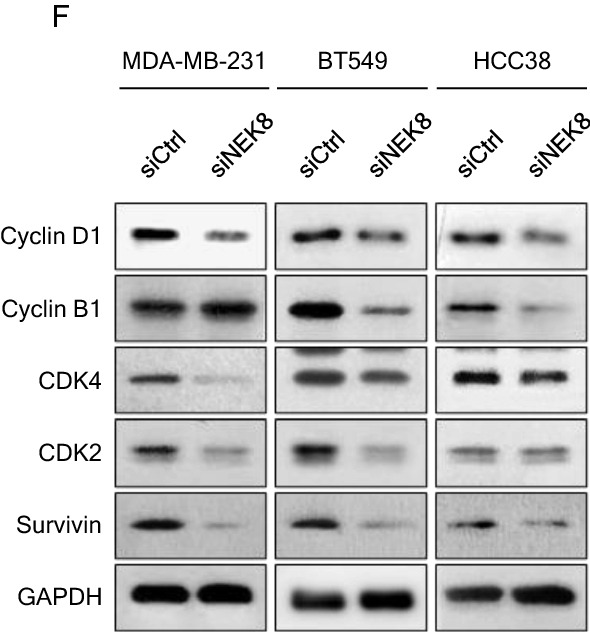
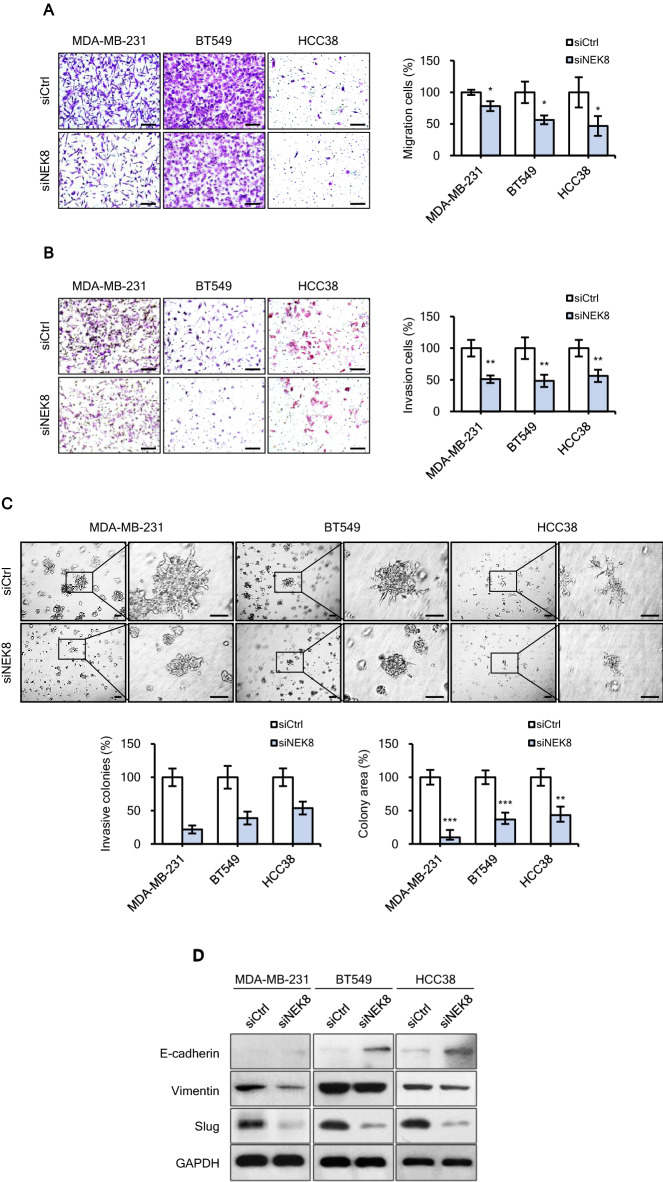
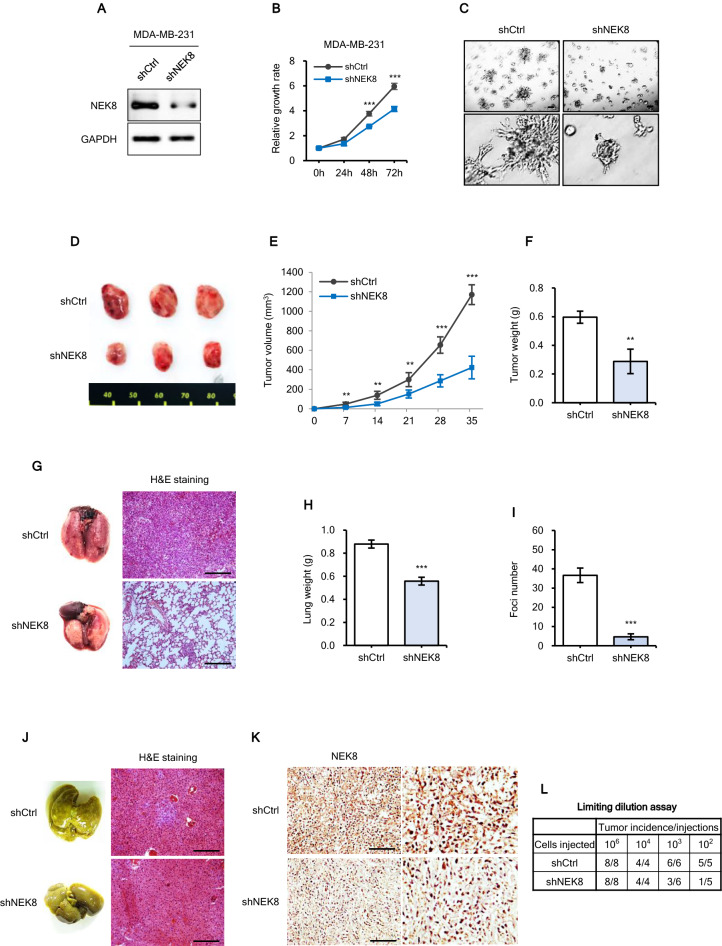
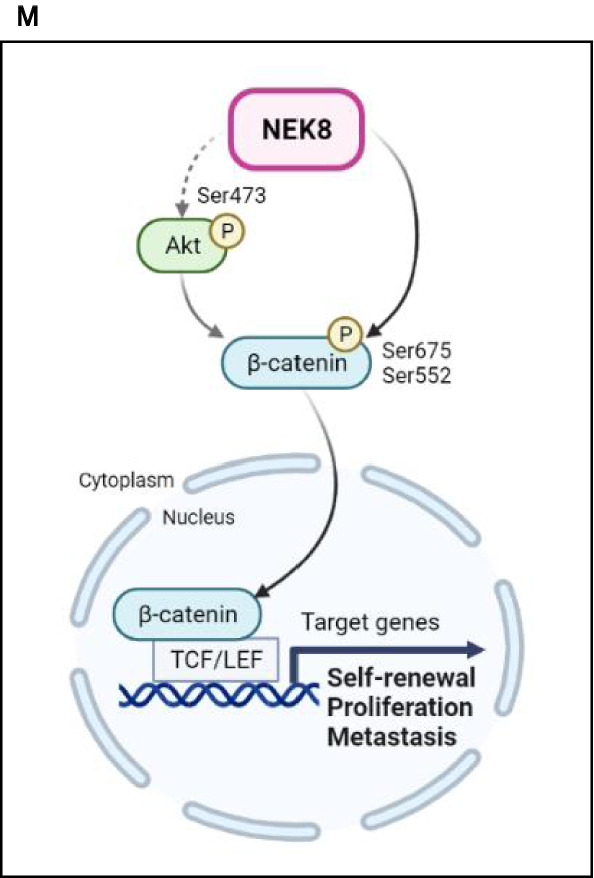
Never in mitosis gene A (NIMA)-related kinase-8 (NEK8) is involved in cell cycle progression, cytoskeleton development, and DNA damage repair. However, its role in breast cancer has not yet been explored. To investigate this, NEK8 was knocked down in MDA-MB-231, BT549, and HCC38 breast cancer cell lines. We observed a decrease in cell proliferation and colony formation owing to regulation of the G1/S and G2/M transitions. Furthermore, the expression of several cell cycle regulatory proteins was altered, including that of cyclin D1, cyclin B1, CDK4, CDK2, and surviving. NEK8 knockdown impaired cell migration and invasion as well as reduced the expression of epithelial-mesenchymal transition markers. Regarding stem-cell characteristics, NEK8 knockdown decreased the tumour sphere formation, aldehyde dehydrogenase activity, and stem-cell marker expression, including that of CD44, Sox2, Oct4a, and Nanog. Further analysis revealed that NEK8 interacts with β-catenin. Also, NEK8 knockdown promoted β-catenin degradation. NEK8-silenced MDA-MB-231 cells inhibited xenograft tumour growth, metastasis, and tumour initiation in vivo. Using the Oncomine and TNMplot public databases, we found a significant correlation between NEK8 overexpression and poor clinical outcomes in breast cancer patients. Thus, NEK8 may be a crucial regulator of breast cancer progression and a potential therapeutic target.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
NEK8, a NIMA-family protein kinase at the core of the ciliary INV complex.Cell Commun Signal. 2025 Apr 7;23(1):170. doi: 10.1186/s12964-025-02143-w. Cell Commun Signal. 2025. PMID: 40189576 Free PMC article. Review.
-
Nek8, a NIMA family kinase member, is overexpressed in primary human breast tumors.Gene. 2004 Mar 17;328:135-42. doi: 10.1016/j.gene.2003.12.002. Gene. 2004. PMID: 15019993
-
Anti-metastasis activity of curcumin against breast cancer via the inhibition of stem cell-like properties and EMT.Phytomedicine. 2019 May;58:152740. doi: 10.1016/j.phymed.2018.11.001. Epub 2018 Nov 12. Phytomedicine. 2019. PMID: 31005718
-
The tumor suppressor pVHL down-regulates never-in-mitosis A-related kinase 8 via hypoxia-inducible factors to maintain cilia in human renal cancer cells.J Biol Chem. 2015 Jan 16;290(3):1389-94. doi: 10.1074/jbc.M114.589226. Epub 2014 Dec 1. J Biol Chem. 2015. PMID: 25451921 Free PMC article.
-
The NEK family of serine/threonine kinases as a biomarker for cancer.Clin Exp Med. 2023 Feb;23(1):17-30. doi: 10.1007/s10238-021-00782-0. Epub 2022 Jan 17. Clin Exp Med. 2023. PMID: 35037094 Review.
Cited by
-
NEK8 regulates colorectal cancer progression via phosphorylating MYC.Cell Commun Signal. 2023 Aug 18;21(1):209. doi: 10.1186/s12964-023-01215-z. Cell Commun Signal. 2023. PMID: 37596667 Free PMC article.
-
The NIMA-related kinase family and cancer.Front Oncol. 2025 Mar 27;15:1556917. doi: 10.3389/fonc.2025.1556917. eCollection 2025. Front Oncol. 2025. PMID: 40212678 Free PMC article. Review.
-
NEK8, a NIMA-family protein kinase at the core of the ciliary INV complex.Cell Commun Signal. 2025 Apr 7;23(1):170. doi: 10.1186/s12964-025-02143-w. Cell Commun Signal. 2025. PMID: 40189576 Free PMC article. Review.
-
NEK kinases in cell cycle regulation, DNA damage response, and cancer progression.Tissue Cell. 2025 Jun;94:102811. doi: 10.1016/j.tice.2025.102811. Epub 2025 Feb 28. Tissue Cell. 2025. PMID: 40037068 Review.
-
Overexpression of the NEK8 kinase inhibits homologous recombination.bioRxiv [Preprint]. 2025 Feb 8:2025.02.07.637121. doi: 10.1101/2025.02.07.637121. bioRxiv. 2025. PMID: 39975112 Free PMC article. Preprint.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
