

The aorta in humans and African great apes, and cardiac output and metabolic levels in human evolution
- PMID: 37100851
- PMCID: PMC10133235
- DOI: 10.1038/s41598-023-33675-1
The aorta in humans and African great apes, and cardiac output and metabolic levels in human evolution
Abstract
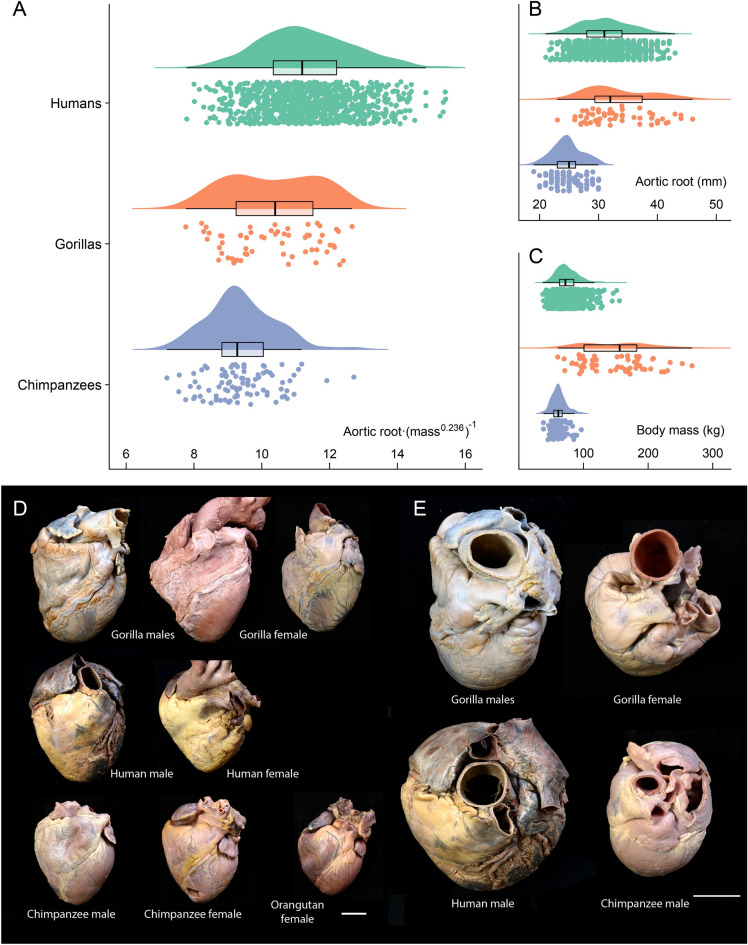
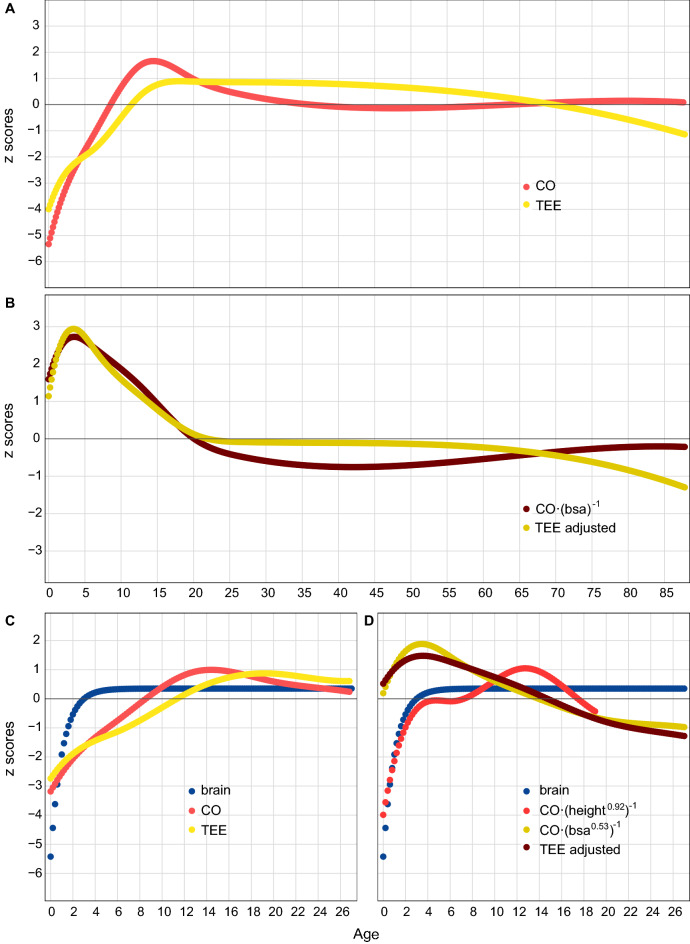
Humans have a larger energy budget than great apes, allowing the combination of the metabolically expensive traits that define our life history. This budget is ultimately related to the cardiac output, the product of the blood pumped from the ventricle and the number of heart beats per minute, a measure of the blood available for the whole organism physiological activity. To show the relationship between cardiac output and energy expenditure in hominid evolution, we study a surrogate measure of cardiac output, the aortic root diameter, in humans and great apes. When compared to gorillas and chimpanzees, humans present an increased body mass adjusted aortic root diameter. We also use data from the literature to show that over the human lifespan, cardiac output and total energy expenditure follow almost identical trajectories, with a marked increase during the period of brain growth, and a plateau during most of the adult life. The limited variation of adjusted cardiac output with sex, age and physical activity supports the compensation model of energy expenditure in humans. Finally, we present a first study of cardiac output in the skeleton through the study of the aortic impression in the vertebral bodies of the spine. It is absent in great apes, and present in humans and Neanderthals, large-brained hominins with an extended life cycle. An increased adjusted cardiac output, underlying higher total energy expenditure, would have been a key process in human evolution.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Analysis of differentially methylated regions in great apes and extinct hominids provides support for the evolutionary hypothesis of schizophrenia.Schizophr Res. 2019 Apr;206:209-216. doi: 10.1016/j.schres.2018.11.025. Epub 2018 Dec 11. Schizophr Res. 2019. PMID: 30545758
-
Patterns of dental development in Homo, Australopithecus, Pan, and Gorilla.Am J Phys Anthropol. 1994 Jul;94(3):307-25. doi: 10.1002/ajpa.1330940303. Am J Phys Anthropol. 1994. PMID: 7943188
-
Do rates of dental wear in extant African great apes inform the time of weaning?J Hum Evol. 2022 Feb;163:103126. doi: 10.1016/j.jhevol.2021.103126. Epub 2021 Dec 23. J Hum Evol. 2022. PMID: 34954400
-
Comparative Pathology of Aging Great Apes: Bonobos, Chimpanzees, Gorillas, and Orangutans.Vet Pathol. 2016 Mar;53(2):250-76. doi: 10.1177/0300985815612154. Epub 2015 Dec 31. Vet Pathol. 2016. PMID: 26721908 Review.
-
Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great ape and human evolution.Proc Natl Acad Sci U S A. 2012 Sep 25;109(39):15716-21. doi: 10.1073/pnas.1211740109. Epub 2012 Aug 13. Proc Natl Acad Sci U S A. 2012. PMID: 22891323 Free PMC article. Review.
Cited by
-
Lack of sex-specific differences in the associations between the dimensions of great vessels and exercise performance in amateur cyclists.PLoS One. 2024 Nov 4;19(11):e0313165. doi: 10.1371/journal.pone.0313165. eCollection 2024. PLoS One. 2024. PMID: 39495753 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
