

Metabololipidomic and proteomic profiling reveals aberrant macrophage activation and interrelated immunomodulatory mediator release during aging
- PMID: 37101405
- PMCID: PMC10352559
- DOI: 10.1111/acel.13856
Metabololipidomic and proteomic profiling reveals aberrant macrophage activation and interrelated immunomodulatory mediator release during aging
Abstract
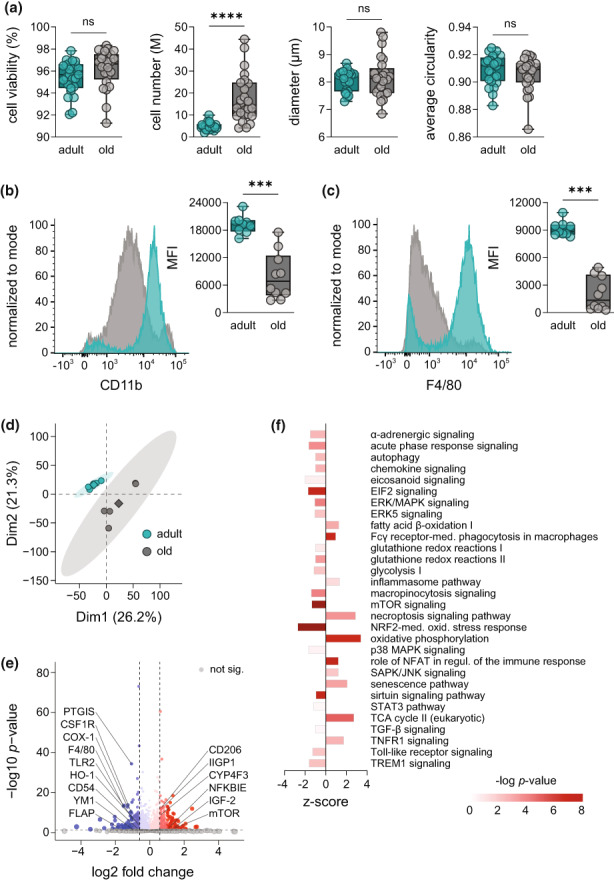
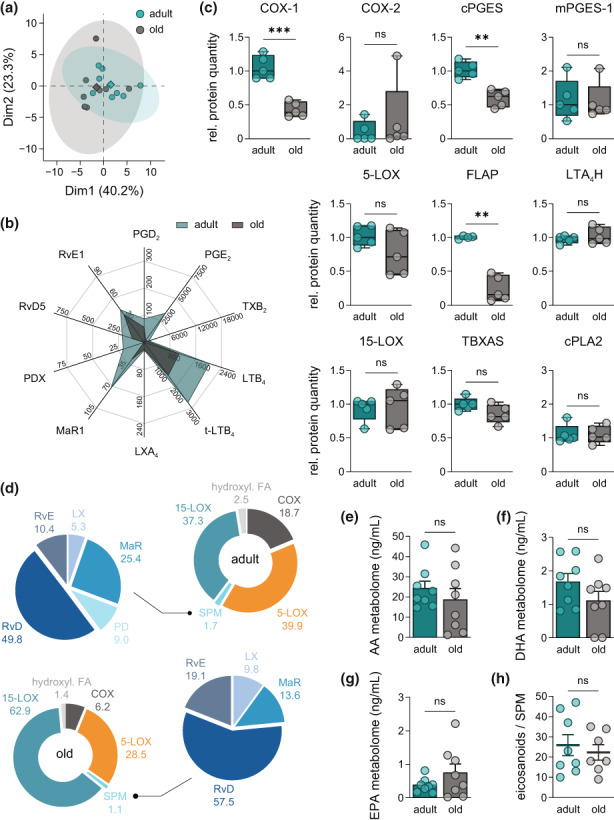
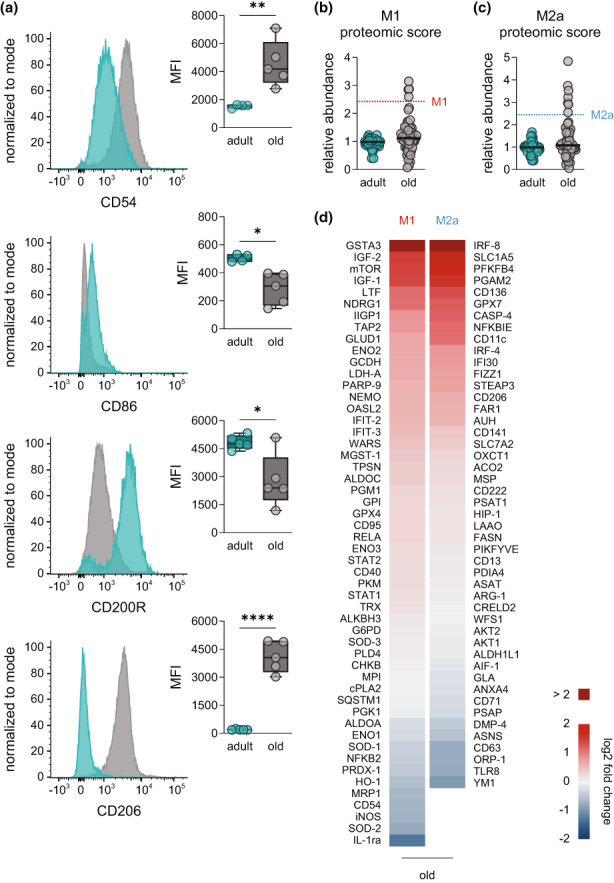
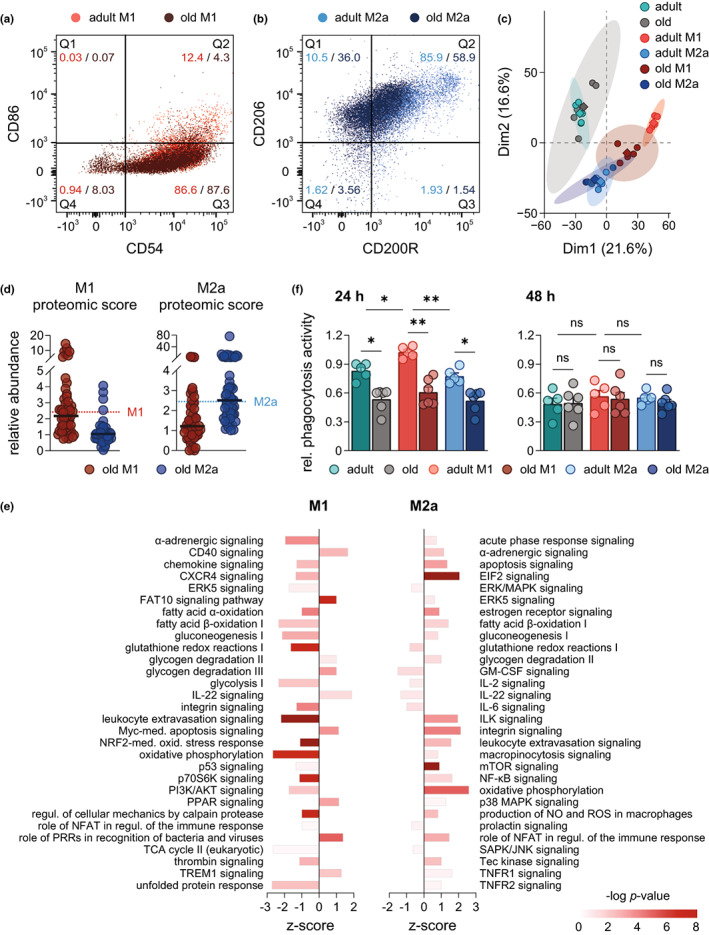
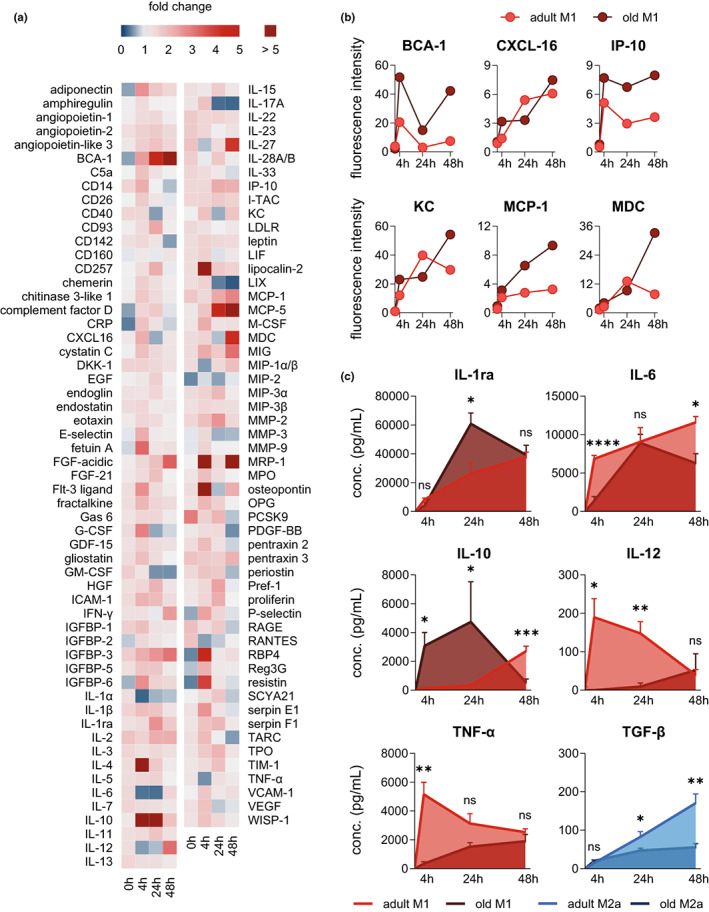
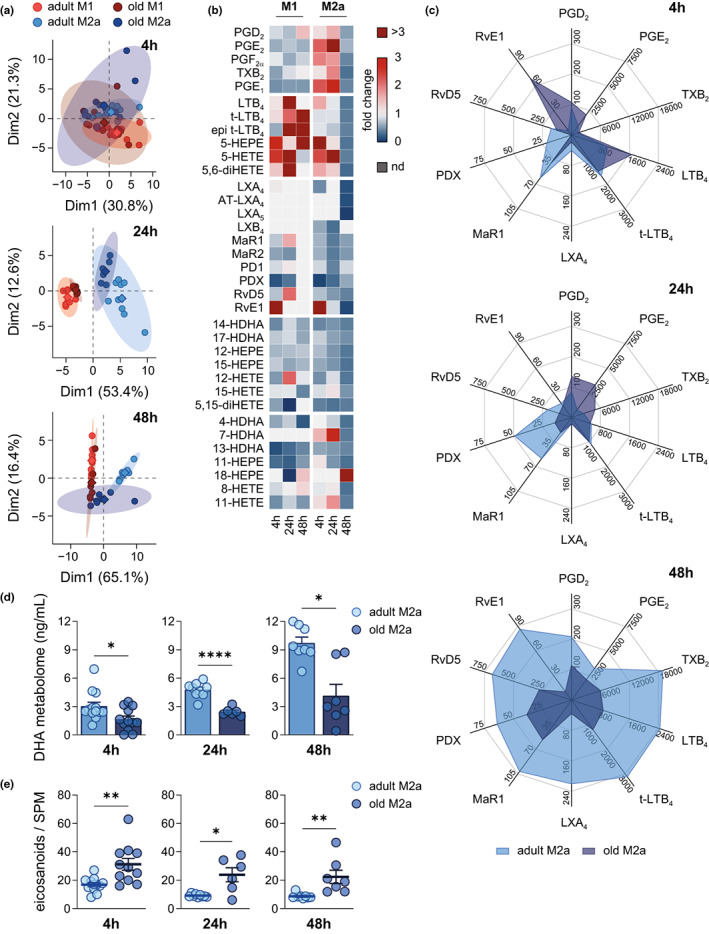
Macrophages adapt distinct pro-inflammatory (M1-like) and pro-resolving (M2-like) phenotypes with specific tasks in the immune response and tissue homeostasis. Altered macrophage responses with age are causative for unresolved inflammation, so-called inflammaging, and lead to higher infection susceptibility with unfavorable progression. Here, we reveal molecular determinants of age-related changes in phenotypic functions of murine peritoneal macrophages (PM) by employing comprehensive mass spectrometry-based proteomics (4746 protein groups) and metabololipidomics (>40 lipid mediators). Divergent expression of various macrophage-specific marker proteins and signaling pathways indicates aberrant PM phenotypes in old mice which detrimentally impact their capabilities to release immunomodulatory chemokines and cytokines. We show that aging strikingly compromises the polarization process of macrophages to adapt either pro-inflammatory or pro-resolving phenotypes, thereby yielding aberrant and afunctional macrophage subtypes that cannot be readily assigned to either a typical M1 or M2 phenotype. In particular, the phenotypic adaptation of the bacteria-challenged metabololipidome in macrophages related to inflammation is severely limited by age, which persists across ex vivo polarization towards M1 and M2a macrophages. Our results establish distinct age-associated PM phenotypes outside of the simplified M1 and M2 dichotomy and challenge the dogma of increased pro-inflammatory macrophage pre-activation due to aging by revealing maladaptive functions throughout all phases of inflammation, including resolution.
Keywords: aging; eicosanoids; inflammation; lipidomics; macrophage activation; mediators of inflammation; peritoneal macrophages; proteomics.
© 2023 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Aiello, A. , Farzaneh, F. , Candore, G. , Caruso, C. , Davinelli, S. , Gambino, C. M. , Ligotti, M. E. , Zareian, N. , & Accardi, G. (2019). Immunosenescence and its hallmarks: How to oppose aging strategically? A review of potential options for therapeutic intervention. Frontiers in Immunology, 10, 2247. 10.3389/fimmu.2019.02247 - DOI - PMC - PubMed
-
- Almanzar, N. , Antony, J. , Baghel, A. S. , Bakerman, I. , Bansal, I. , Barres, B. A. , Beachy, P. A. , Berdnik, D. , Bilen, B. , Brownfield, D. , Cain, C. , Chan, C. K. F. , Chen, M. B. , Clarke, M. F. , Conley, S. D. , Darmanis, S. , Demers, A. , Demir, K. , De Morree, A. , … Zou, J. (2020). A single‐cell transcriptomic atlas characterizes ageing tissues in the mouse. Nature, 583(7817), 590–595. 10.1038/s41586-020-2496-1 - DOI - PMC - PubMed
-
- Arnardottir, H. H. , Dalli, J. , Colas, R. A. , Shinohara, M. , & Serhan, C. N. (2014). Aging delays resolution of acute inflammation in mice: Reprogramming the host response with novel nano‐proresolving medicines. The Journal of Immunology, 193(8), 4235–4244. 10.4049/jimmunol.1401313 - DOI - PMC - PubMed
-
- Bain, C. C. , Hawley, C. A. , Garner, H. , Scott, C. L. , Schridde, A. , Steers, N. J. , Mack, M. , Joshi, A. , Guilliams, M. , Mowat, A. M. I. , Geissmann, F. , & Jenkins, S. J. (2016). Long‐lived self‐renewing bone marrow‐derived macrophages displace embryo‐derived cells to inhabit adult serous cavities. Nature Communications, 7(1), 11852. 10.1038/ncomms11852 - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
