

Inhibition of 3-Hydroxykynurenine Transaminase from Aedes aegypti and Anopheles gambiae: A Mosquito-Specific Target to Combat the Transmission of Arboviruses
- PMID: 37101811
- PMCID: PMC10125267
- DOI: 10.1021/acsbiomedchemau.2c00080
Inhibition of 3-Hydroxykynurenine Transaminase from Aedes aegypti and Anopheles gambiae: A Mosquito-Specific Target to Combat the Transmission of Arboviruses
Abstract
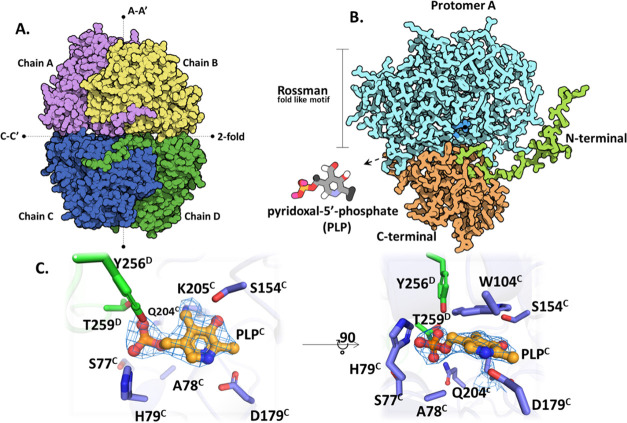
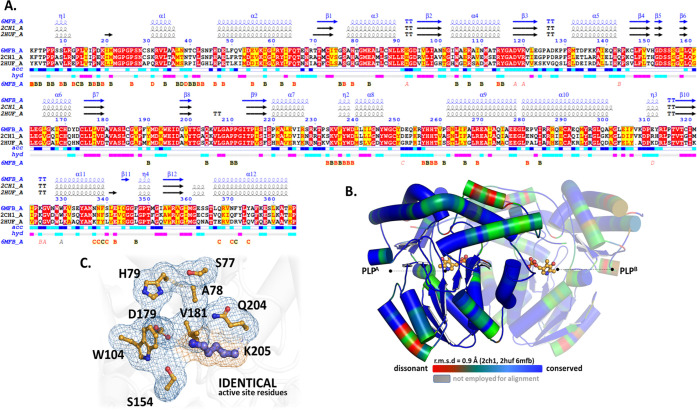
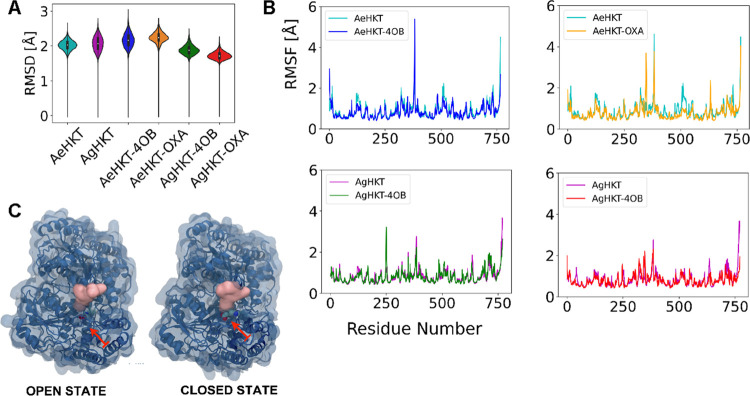
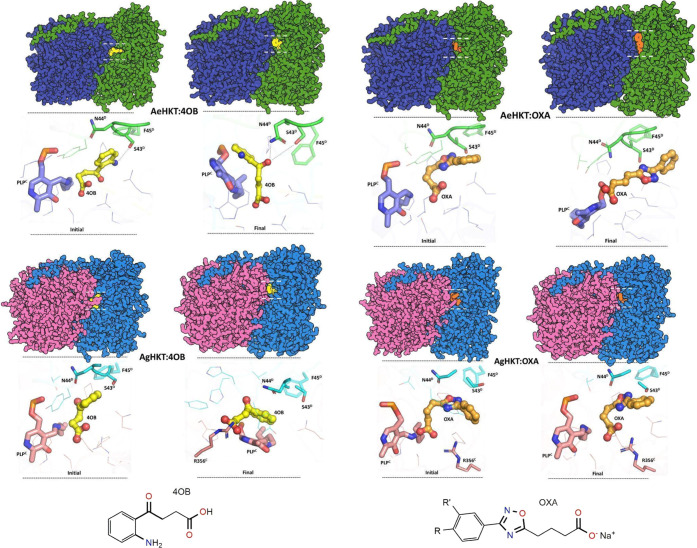
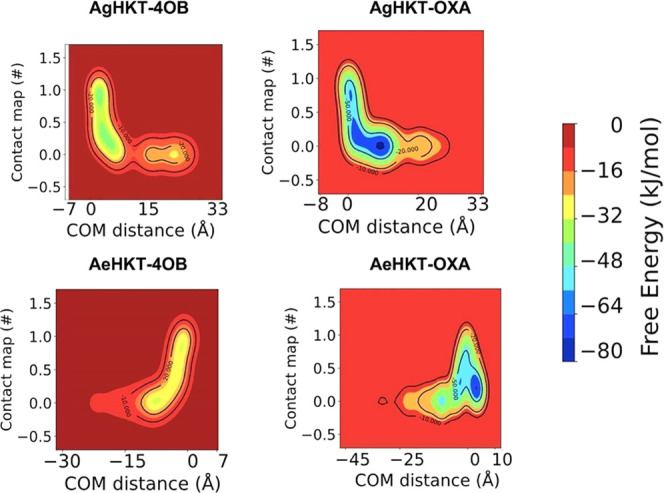
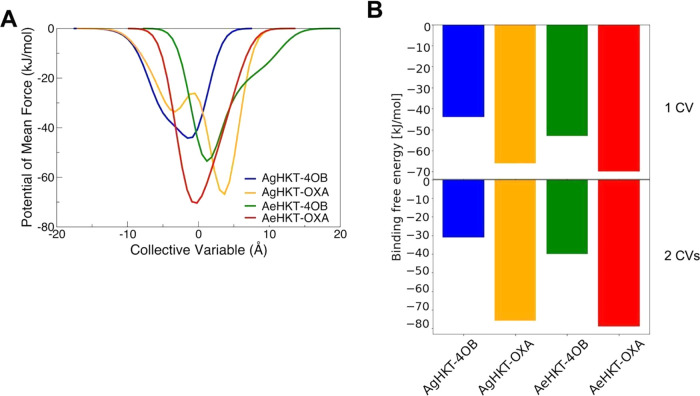
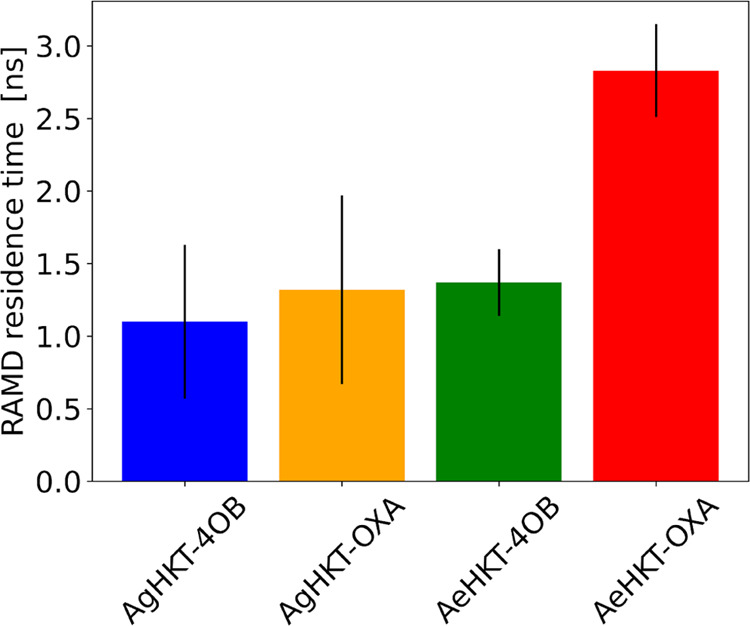
Arboviral infections such as Zika, chikungunya, dengue, and yellow fever pose significant health problems globally. The population at risk is expanding with the geographical distribution of the main transmission vector of these viruses, the Aedes aegypti mosquito. The global spreading of this mosquito is driven by human migration, urbanization, climate change, and the ecological plasticity of the species. Currently, there are no specific treatments for Aedes-borne infections. One strategy to combat different mosquito-borne arboviruses is to design molecules that can specifically inhibit a critical host protein. We obtained the crystal structure of 3-hydroxykynurenine transaminase (AeHKT) from A. aegypti, an essential detoxification enzyme of the tryptophan metabolism pathway. Since AeHKT is found exclusively in mosquitoes, it provides the ideal molecular target for the development of inhibitors. Therefore, we determined and compared the free binding energy of the inhibitors 4-(2-aminophenyl)-4-oxobutyric acid (4OB) and sodium 4-(3-phenyl-1,2,4-oxadiazol-5-yl)butanoate (OXA) to AeHKT and AgHKT from Anopheles gambiae, the only crystal structure of this enzyme previously known. The cocrystallized inhibitor 4OB binds to AgHKT with K i of 300 μM. We showed that OXA binds to both AeHKT and AgHKT enzymes with binding energies 2-fold more favorable than the crystallographic inhibitor 4OB and displayed a 2-fold greater residence time τ upon binding to AeHKT than 4OB. These findings indicate that the 1,2,4-oxadiazole derivatives are inhibitors of the HKT enzyme not only from A. aegypti but also from A. gambiae.
© 2023 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
A second generation of 1,2,4-oxadiazole derivatives with enhanced solubility for inhibition of 3-hydroxykynurenine transaminase (HKT) from Aedes aegypti.RSC Med Chem. 2020 Dec 9;12(2):222-236. doi: 10.1039/d0md00305k. eCollection 2021 Mar 4. RSC Med Chem. 2020. PMID: 34046611 Free PMC article.
-
Discovery of 1,2,4-oxadiazole derivatives as a novel class of noncompetitive inhibitors of 3-hydroxykynurenine transaminase (HKT) from Aedes aegypti.Bioorg Med Chem. 2020 Jan 15;28(2):115252. doi: 10.1016/j.bmc.2019.115252. Epub 2019 Dec 9. Bioorg Med Chem. 2020. PMID: 31864777
-
The enzyme 3-hydroxykynurenine transaminase as potential target for 1,2,4-oxadiazoles with larvicide activity against the dengue vector Aedes aegypti.Bioorg Med Chem. 2013 Nov 15;21(22):6996-7003. doi: 10.1016/j.bmc.2013.09.020. Epub 2013 Sep 20. Bioorg Med Chem. 2013. PMID: 24095017
-
Minireview: Epidemiological impact of arboviral diseases in Latin American countries, arbovirus-vector interactions and control strategies.Pathog Dis. 2021 Sep 6;79(7):ftab043. doi: 10.1093/femspd/ftab043. Pathog Dis. 2021. PMID: 34410378 Review.
-
Globe-Trotting Aedes aegypti and Aedes albopictus: Risk Factors for Arbovirus Pandemics.Vector Borne Zoonotic Dis. 2020 Feb;20(2):71-81. doi: 10.1089/vbz.2019.2486. Epub 2019 Sep 26. Vector Borne Zoonotic Dis. 2020. PMID: 31556813 Free PMC article. Review.
Cited by
-
Exploring the Larvicidal and Adulticidal Activity against Aedes aegypti of Essential Oil from Bocageopsis multiflora.Molecules. 2024 May 10;29(10):2240. doi: 10.3390/molecules29102240. Molecules. 2024. PMID: 38792102 Free PMC article.
-
Molecular docking and molecular dynamics simulation studies of inhibitor candidates against Anopheles gambiae 3-hydroxykynurenine transaminase and implications on vector control.Heliyon. 2025 Jan 2;11(1):e41633. doi: 10.1016/j.heliyon.2025.e41633. eCollection 2025 Jan 15. Heliyon. 2025. PMID: 39866405 Free PMC article.
References
-
- Brady O. J.; Gething P. W.; Bhatt S.; Messina J. P.; Brownstein J. S.; Hoen A. G.; Moyes C. L.; Farlow A. W.; Scott T. W.; Hay S. I. Refining the Global Spatial Limits of Dengue Virus Transmission by Evidence-Based Consensus. PLoS Negl. Trop. Dis. 2012, 6, e176010.1371/journal.pntd.0001760. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
