Relating enhancer genetic variation across mammals to complex phenotypes using machine learning
- PMID: 37104615
- PMCID: PMC10322212
- DOI: 10.1126/science.abm7993
Relating enhancer genetic variation across mammals to complex phenotypes using machine learning
Abstract
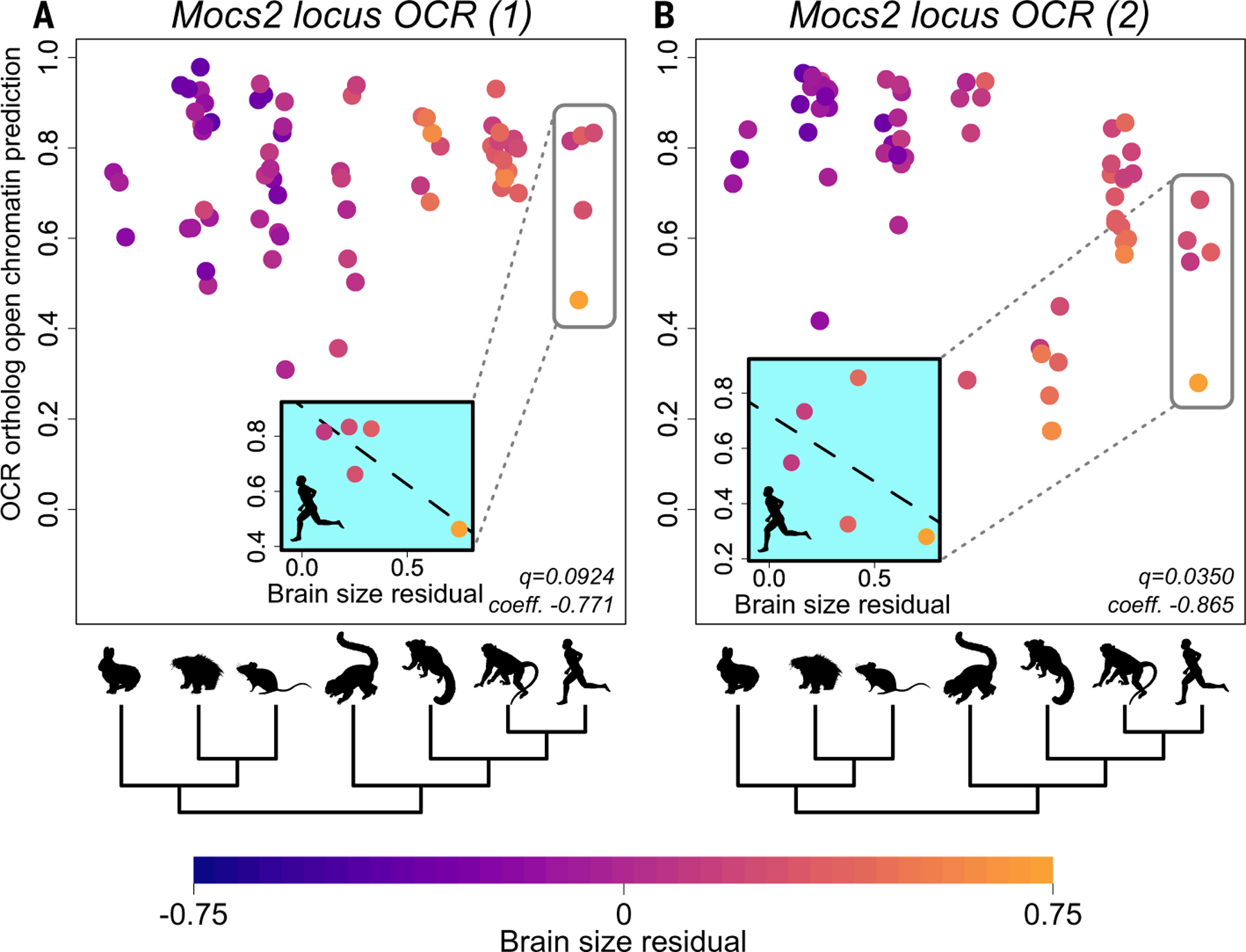
Protein-coding differences between species often fail to explain phenotypic diversity, suggesting the involvement of genomic elements that regulate gene expression such as enhancers. Identifying associations between enhancers and phenotypes is challenging because enhancer activity can be tissue-dependent and functionally conserved despite low sequence conservation. We developed the Tissue-Aware Conservation Inference Toolkit (TACIT) to associate candidate enhancers with species' phenotypes using predictions from machine learning models trained on specific tissues. Applying TACIT to associate motor cortex and parvalbumin-positive interneuron enhancers with neurological phenotypes revealed dozens of enhancer-phenotype associations, including brain size-associated enhancers that interact with genes implicated in microcephaly or macrocephaly. TACIT provides a foundation for identifying enhancers associated with the evolution of any convergently evolved phenotype in any large group of species with aligned genomes.
Conflict of interest statement
Figures