

Regulation of phospholipid distribution in the lipid bilayer by flippases and scramblases
- PMID: 37106071
- PMCID: PMC10134735
- DOI: 10.1038/s41580-023-00604-z
Regulation of phospholipid distribution in the lipid bilayer by flippases and scramblases
Erratum in
-
Publisher Correction: Regulation of phospholipid distribution in the lipid bilayer by flippases and scramblases.Nat Rev Mol Cell Biol. 2023 Aug;24(8):597. doi: 10.1038/s41580-023-00621-y. Nat Rev Mol Cell Biol. 2023. PMID: 37217623 Free PMC article. No abstract available.
Abstract
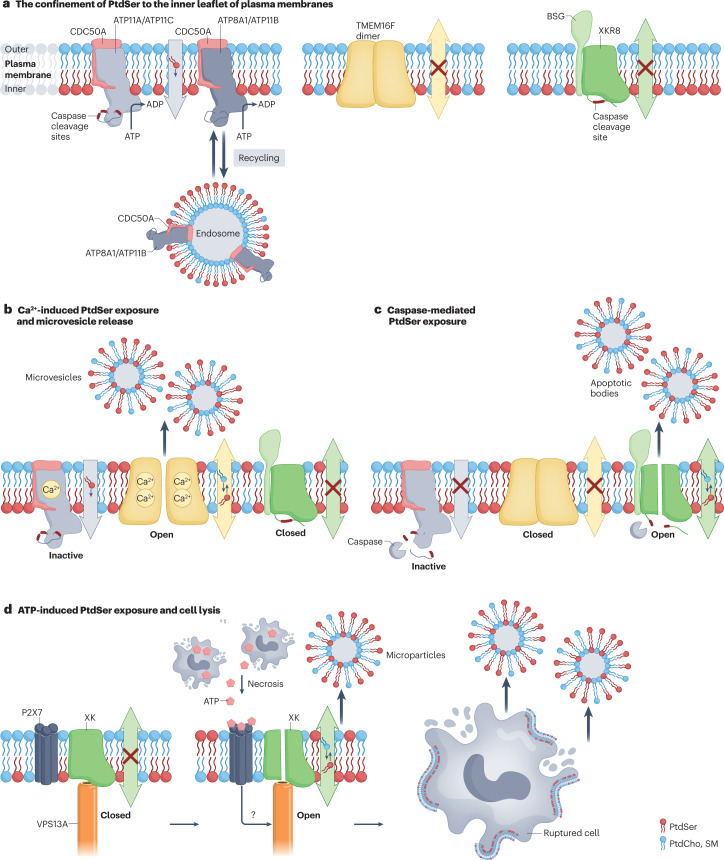
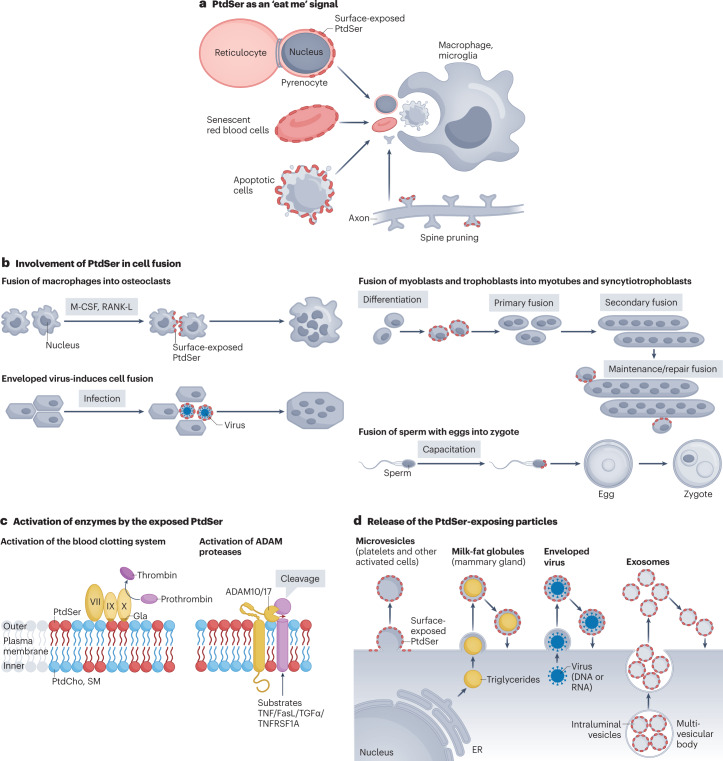
Cellular membranes function as permeability barriers that separate cells from the external environment or partition cells into distinct compartments. These membranes are lipid bilayers composed of glycerophospholipids, sphingolipids and cholesterol, in which proteins are embedded. Glycerophospholipids and sphingolipids freely move laterally, whereas transverse movement between lipid bilayers is limited. Phospholipids are asymmetrically distributed between membrane leaflets but change their location in biological processes, serving as signalling molecules or enzyme activators. Designated proteins - flippases and scramblases - mediate this lipid movement between the bilayers. Flippases mediate the confined localization of specific phospholipids (phosphatidylserine (PtdSer) and phosphatidylethanolamine) to the cytoplasmic leaflet. Scramblases randomly scramble phospholipids between leaflets and facilitate the exposure of PtdSer on the cell surface, which serves as an important signalling molecule and as an 'eat me' signal for phagocytes. Defects in flippases and scramblases cause various human diseases. We herein review the recent research on the structure of flippases and scramblases and their physiological roles. Although still poorly understood, we address the mechanisms by which they translocate phospholipids between lipid bilayers and how defects cause human diseases.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Murate M, et al. Transbilayer distribution of lipids at nano scale. J. Cell Sci. 2015;128:1627–1638. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
