

The Interplay between Intracellular Iron Homeostasis and Neuroinflammation in Neurodegenerative Diseases
- PMID: 37107292
- PMCID: PMC10135822
- DOI: 10.3390/antiox12040918
The Interplay between Intracellular Iron Homeostasis and Neuroinflammation in Neurodegenerative Diseases
Abstract
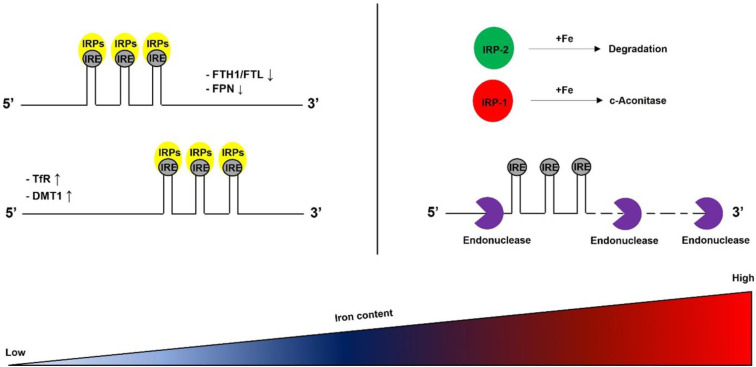
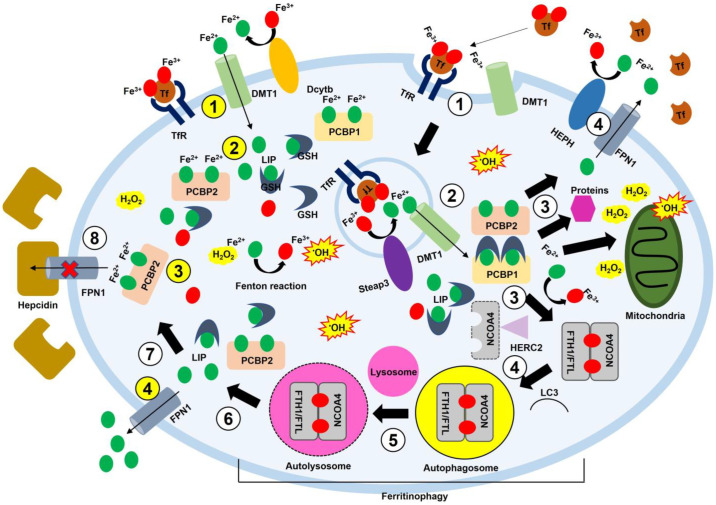
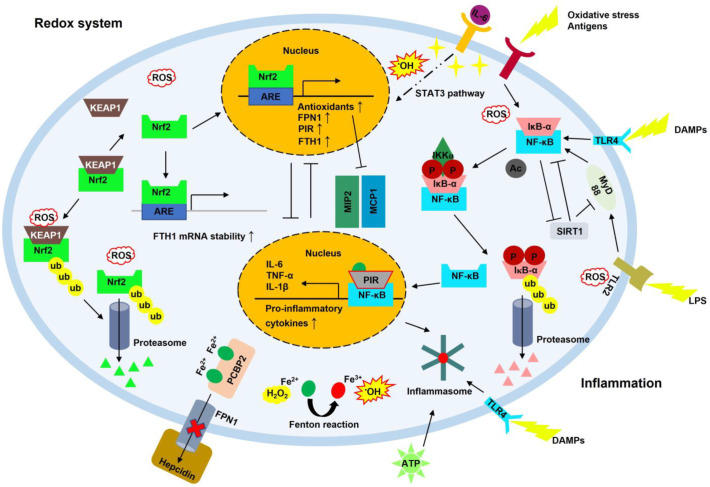
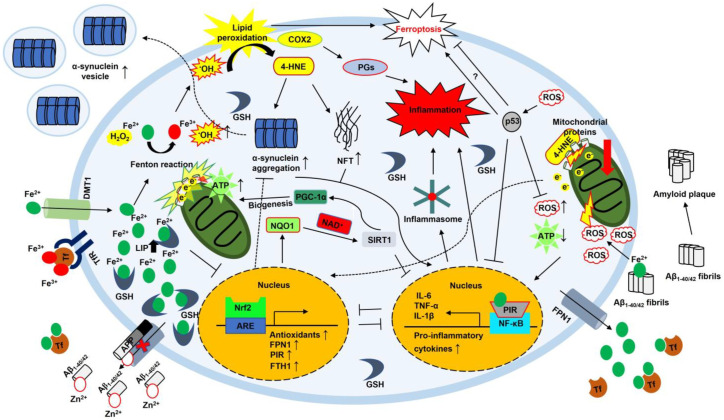
Iron is essential for life. Many enzymes require iron for appropriate function. However, dysregulation of intracellular iron homeostasis produces excessive reactive oxygen species (ROS) via the Fenton reaction and causes devastating effects on cells, leading to ferroptosis, an iron-dependent cell death. In order to protect against harmful effects, the intracellular system regulates cellular iron levels through iron regulatory mechanisms, including hepcidin-ferroportin, divalent metal transporter 1 (DMT1)-transferrin, and ferritin-nuclear receptor coactivator 4 (NCOA4). During iron deficiency, DMT1-transferrin and ferritin-NCOA4 systems increase intracellular iron levels via endosomes and ferritinophagy, respectively. In contrast, repleting extracellular iron promotes cellular iron absorption through the hepcidin-ferroportin axis. These processes are regulated by the iron-regulatory protein (IRP)/iron-responsive element (IRE) system and nuclear factor erythroid 2-related factor 2 (Nrf2). Meanwhile, excessive ROS also promotes neuroinflammation by activating the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB). NF-κB forms inflammasomes, inhibits silent information regulator 2-related enzyme 1 (SIRT1), and induces pro-inflammatory cytokines (IL-6, TNF-α, and IL-1β). Furthermore, 4-hydroxy-2,3-trans-nonenal (4-HNE), the end-product of ferroptosis, promotes the inflammatory response by producing amyloid-beta (Aβ) fibrils and neurofibrillary tangles in Alzheimer's disease, and alpha-synuclein aggregation in Parkinson's disease. This interplay shows that intracellular iron homeostasis is vital to maintain inflammatory homeostasis. Here, we review the role of iron homeostasis in inflammation based on recent findings.
Keywords: 4-HNE; NF-κB; Nrf2; ferroptosis; intracellular iron homeostasis; neurodegenerative diseases; neuroinflammation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
