

Early Steps towards Hearing: Placodes and Sensory Development
- PMID: 37108158
- PMCID: PMC10139157
- DOI: 10.3390/ijms24086994
Early Steps towards Hearing: Placodes and Sensory Development
Abstract
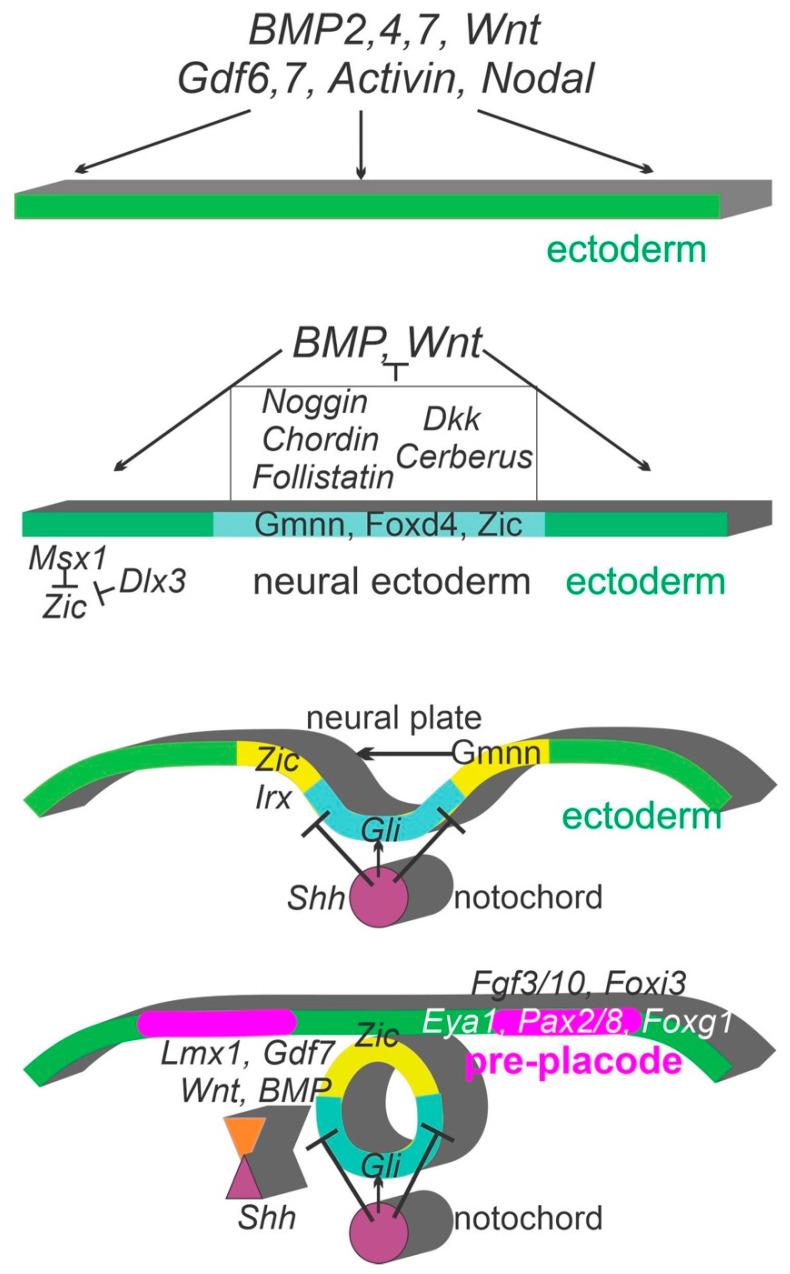
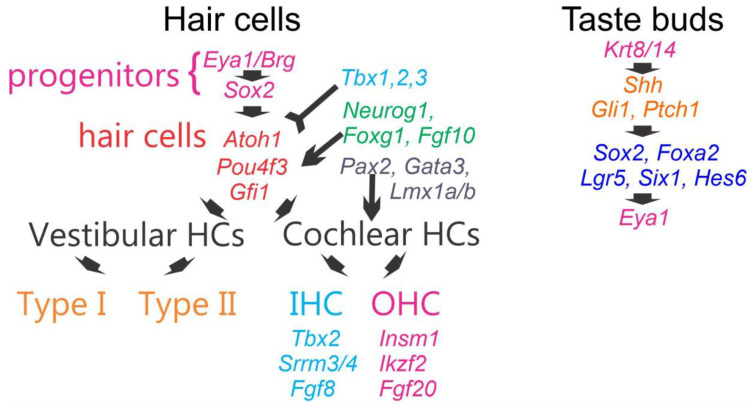
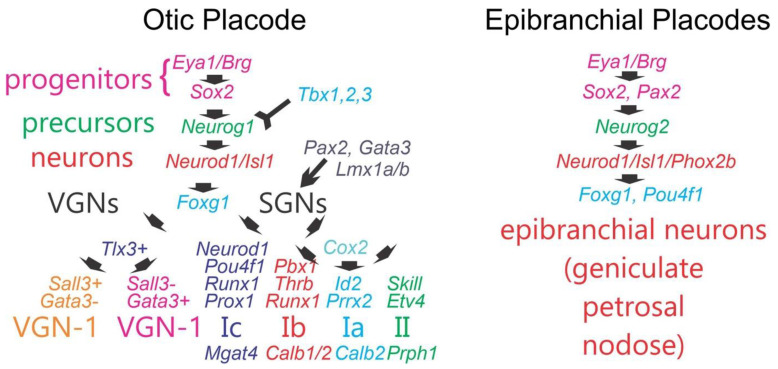
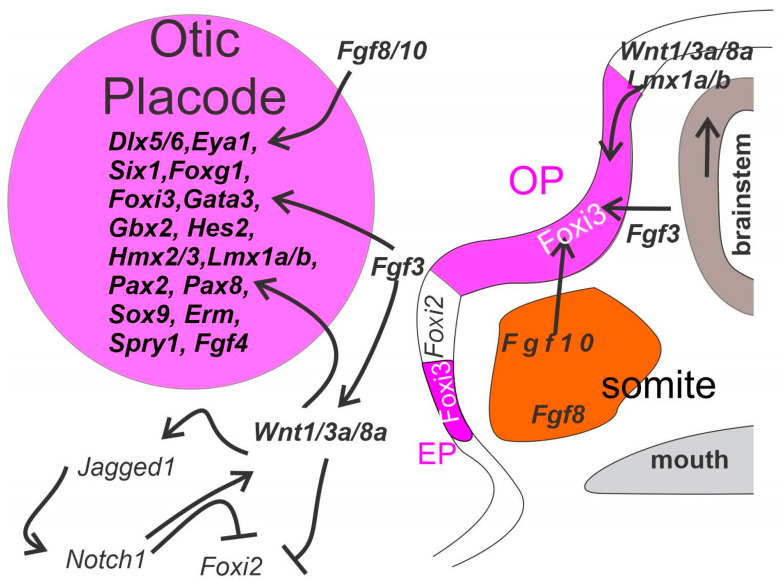
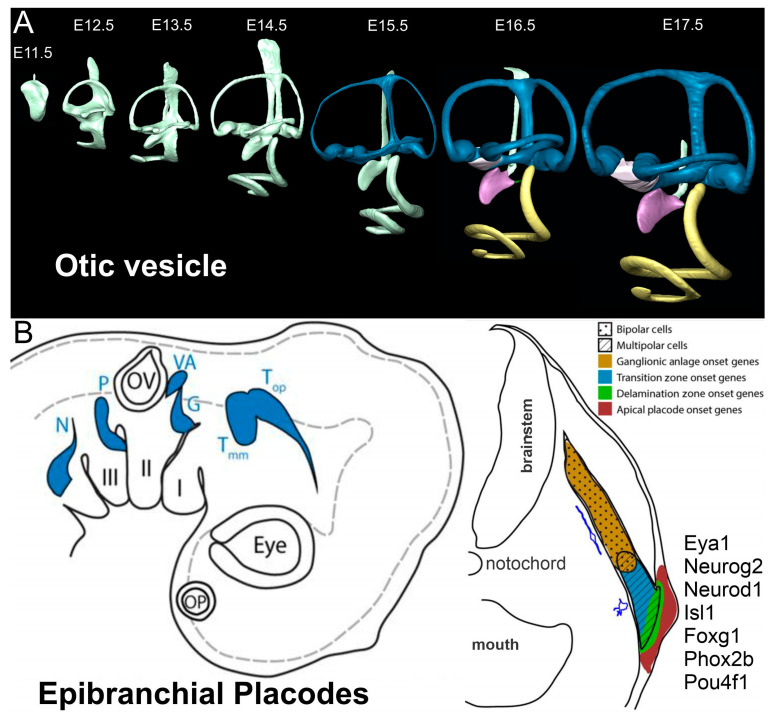
Sensorineural hearing loss is the most prevalent sensory deficit in humans. Most cases of hearing loss are due to the degeneration of key structures of the sensory pathway in the cochlea, such as the sensory hair cells, the primary auditory neurons, and their synaptic connection to the hair cells. Different cell-based strategies to replace damaged inner ear neurosensory tissue aiming at the restoration of regeneration or functional recovery are currently the subject of intensive research. Most of these cell-based treatment approaches require experimental in vitro models that rely on a fine understanding of the earliest morphogenetic steps that underlie the in vivo development of the inner ear since its initial induction from a common otic-epibranchial territory. This knowledge will be applied to various proposed experimental cell replacement strategies to either address the feasibility or identify novel therapeutic options for sensorineural hearing loss. In this review, we describe how ear and epibranchial placode development can be recapitulated by focusing on the cellular transformations that occur as the inner ear is converted from a thickening of the surface ectoderm next to the hindbrain known as the otic placode to an otocyst embedded in the head mesenchyme. Finally, we will highlight otic and epibranchial placode development and morphogenetic events towards progenitors of the inner ear and their neurosensory cell derivatives.
Keywords: epibranchial; gene regulatory network; hair cell; inner ear; sensory neuron; sensory placode; taste buds.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Early steps in inner ear development: induction and morphogenesis of the otic placode.Front Pharmacol. 2015 Feb 10;6:19. doi: 10.3389/fphar.2015.00019. eCollection 2015. Front Pharmacol. 2015. PMID: 25713536 Free PMC article. Review.
-
Zebrafish Foxi1 provides a neuronal ground state during inner ear induction preceding the Dlx3b/4b-regulated sensory lineage.Development. 2013 May;140(9):1936-45. doi: 10.1242/dev.087718. Development. 2013. PMID: 23571216
-
Establishment of mice expressing EGFP in the placode-derived inner ear sensory cell lineage and FACS-array analysis focused on the regional specificity of the otocyst.J Comp Neurol. 2010 Dec 1;518(23):4702-22. doi: 10.1002/cne.22468. J Comp Neurol. 2010. PMID: 20963824
-
Growth factor therapy to the damaged inner ear: clinical prospects.Int J Pediatr Otorhinolaryngol. 1999 Oct 5;49 Suppl 1:S19-25. doi: 10.1016/s0165-5876(99)00126-3. Int J Pediatr Otorhinolaryngol. 1999. PMID: 10577769 Review.
-
Fgf8 and Fgf3 are required for zebrafish ear placode induction, maintenance and inner ear patterning.Mech Dev. 2002 Nov;119(1):91-108. doi: 10.1016/s0925-4773(02)00343-x. Mech Dev. 2002. PMID: 12385757
Cited by
-
Shared and organ-specific gene-expression programs during the development of the cochlea and the superior olivary complex.RNA Biol. 2023 Jan;20(1):629-640. doi: 10.1080/15476286.2023.2247628. RNA Biol. 2023. PMID: 37602850 Free PMC article.
-
Placode and neural crest origins of congenital deafness in mouse models of Waardenburg-Shah syndrome.iScience. 2024 Dec 24;28(1):111680. doi: 10.1016/j.isci.2024.111680. eCollection 2025 Jan 17. iScience. 2024. PMID: 39868048 Free PMC article.
-
Timing and Graded BMP Signalling Determines Fate of Neural Crest and Ectodermal Placode Derivatives from Pluripotent Stem Cells.Biomedicines. 2024 Oct 4;12(10):2262. doi: 10.3390/biomedicines12102262. Biomedicines. 2024. PMID: 39457575 Free PMC article. Review.
-
Research progress on the treatment and nursing of sensorineural hearing loss.Front Neurosci. 2023 Jun 6;17:1199946. doi: 10.3389/fnins.2023.1199946. eCollection 2023. Front Neurosci. 2023. PMID: 37346087 Free PMC article. Review.
-
Development of ectodermal and endodermal taste buds.Dev Biol. 2025 Feb;518:20-27. doi: 10.1016/j.ydbio.2024.10.005. Epub 2024 Oct 30. Dev Biol. 2025. PMID: 39486632 Review.
References
-
- Xu J., Li J., Zhang T., Jiang H., Ramakrishnan A., Fritzsch B., Shen L., Xu P.X. Chromatin remodelers and lineage-specific factors interact to target enhancers to establish proneurosensory fate within otic ectoderm. Proc. Natl. Acad. Sci. USA. 2021;118:1–12. doi: 10.1073/pnas.2025196118. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
