

The Thioredoxin System in Edwardsiella piscicida Contributes to Oxidative Stress Tolerance, Motility, and Virulence
- PMID: 37110252
- PMCID: PMC10145099
- DOI: 10.3390/microorganisms11040827
The Thioredoxin System in Edwardsiella piscicida Contributes to Oxidative Stress Tolerance, Motility, and Virulence
Abstract
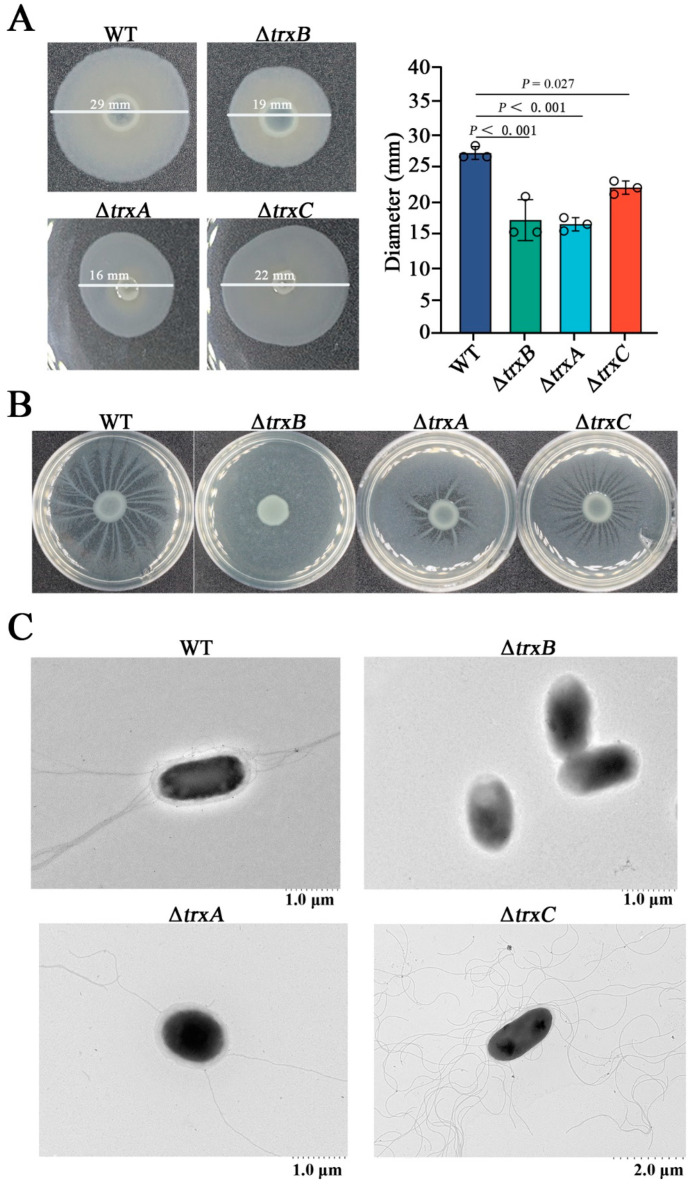
Edwardsiella piscicida is an important fish pathogen that causes substantial economic losses. In order to understand its pathogenic mechanism, additional new virulence factors need to be identified. The bacterial thioredoxin system is a major disulfide reductase system, but its function is largely unknown in E. piscicida. In this study, we investigated the roles of the thioredoxin system in E. piscicida (named TrxBEp, TrxAEp, and TrxCEp, respectively) by constructing a correspondingly markerless in-frame mutant strain: ΔtrxB, ΔtrxA, and ΔtrxC, respectively. We found that (i) TrxBEp is confirmed as an intracellular protein, which is different from the prediction made by the Protter illustration; (ii) compared to the wild-type strain, ΔtrxB exhibits resistance against H2O2 stress but high sensitivity to thiol-specific diamide stress, while ΔtrxA and ΔtrxC are moderately sensitive to both H2O2 and diamide conditions; (iii) the deletions of trxBEp, trxAEp, and trxCEp damage E. piscicida's flagella formation and motility, and trxBEp plays a decisive role; (iv) deletions of trxBEp, trxAEp, and trxCEp substantially abate bacterial resistance against host serum, especially trxBEp deletion; (v) trxAEp and trxCEp, but not trxBEp, are involved in bacterial survival and replication in phagocytes; (vi) the thioredoxin system participates in bacterial dissemination in host immune tissues. These findings indicate that the thioredoxin system of E. piscicida plays an important role in stress resistance and virulence, which provides insight into the pathogenic mechanism of E. piscicida.
Keywords: Edwardsiella piscicida; motility; oxidative stress; thioredoxin system; virulence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







Similar articles
-
Thioredoxin H (TrxH) contributes to adversity adaptation and pathogenicity of Edwardsiella piscicida.Vet Res. 2019 Apr 15;50(1):26. doi: 10.1186/s13567-019-0645-z. Vet Res. 2019. PMID: 30992061 Free PMC article.
-
Universal stress proteins contribute Edwardsiella piscicida adversity resistance and pathogenicity and promote blocking host immune response.Fish Shellfish Immunol. 2019 Dec;95:248-258. doi: 10.1016/j.fsi.2019.10.035. Epub 2019 Oct 22. Fish Shellfish Immunol. 2019. PMID: 31654767
-
Edwardsiella piscicida Interferes with Classical Endocytic Trafficking and Replicates in a Specialized Replication-Permissive Niche in Nonphagocytic Cells.J Bacteriol. 2021 Jul 22;203(16):e0050520. doi: 10.1128/JB.00505-20. Epub 2021 Jul 22. J Bacteriol. 2021. PMID: 34060905 Free PMC article.
-
Edwardsiella piscicida: A versatile emerging pathogen of fish.Virulence. 2019 Dec;10(1):555-567. doi: 10.1080/21505594.2019.1621648. Virulence. 2019. PMID: 31122125 Free PMC article. Review.
-
Edwardsiella piscicida: a significant bacterial pathogen of cultured fish.Dis Aquat Organ. 2018 Oct 16;131(1):59-71. doi: 10.3354/dao03281. Dis Aquat Organ. 2018. PMID: 30324915 Review.
Cited by
-
Temperature-dependent alterations in the proteome of the emergent fish pathogen Edwardsiella piscicida.J Fish Dis. 2025 Sep;48(9):e14017. doi: 10.1111/jfd.14017. Epub 2024 Sep 20. J Fish Dis. 2025. PMID: 39304982 Free PMC article.
-
Comparative genome analyses reveal insights into the marine adaptation of a novel bacterium Pontimicrobium maritimus sp. nov., isolated from seawater.Sci Rep. 2025 Jul 5;15(1):24027. doi: 10.1038/s41598-025-07372-0. Sci Rep. 2025. PMID: 40617894 Free PMC article.
References
-
- Hoshina T. On a new bacterium, Paracolobactrum anguillimortiferum n. sp. Bull. Jpn. Soc. Sci. Fish. 1962;28:162–164. doi: 10.2331/suisan.28.162. - DOI
-
- Ewing W.H., Mcwhorter A.C., Escobar M.R., Lubin A.H. Edwardsiella, a new genus of Enterobacteriaceae based on a new species, E. tarda. Int. Bull. Bacteriol. Nomencl. Taxon. 1965;15:33–38. doi: 10.1099/00207713-15-1-33. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
