

Establishment of an Antiplasmodial Vaccine Based on PfRH5-Encoding RNA Replicons Stabilized by Cationic Liposomes
- PMID: 37111706
- PMCID: PMC10145066
- DOI: 10.3390/pharmaceutics15041223
Establishment of an Antiplasmodial Vaccine Based on PfRH5-Encoding RNA Replicons Stabilized by Cationic Liposomes
Abstract
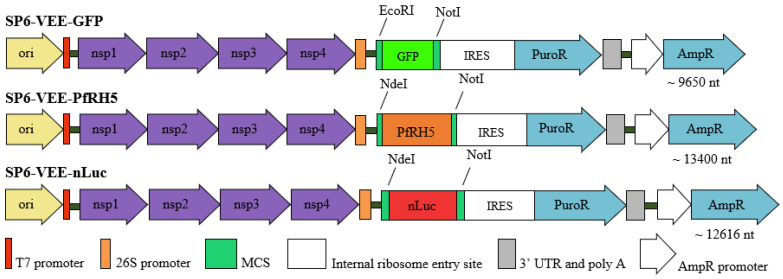
Background: Nucleic acid-based vaccines have been studied for the past four decades, but the approval of the first messenger RNA (mRNA) vaccines during the COVID-19 pandemic opened renewed perspectives for the development of similar vaccines against different infectious diseases. Presently available mRNA vaccines are based on non-replicative mRNA, which contains modified nucleosides encased in lipid vesicles, allowing for entry into the host cell cytoplasm, and reducing inflammatory reactions. An alternative immunization strategy employs self-amplifying mRNA (samRNA) derived from alphaviruses, but lacks viral structural genes. Once incorporated into ionizable lipid shells, these vaccines lead to enhanced gene expression, and lower mRNA doses are required to induce protective immune responses. In the present study, we tested a samRNA vaccine formulation based on the SP6 Venezuelan equine encephalitis (VEE) vector incorporated into cationic liposomes (dimethyldioctadecyl ammonium bromide and a cholesterol derivative). Three vaccines were generated that encoded two reporter genes (GFP and nanoLuc) and the Plasmodium falciparum reticulocyte binding protein homologue 5 (PfRH5).
Methods: Transfection assays were performed using Vero and HEK293T cells, and the mice were immunized via the intradermal route using a tattooing device.
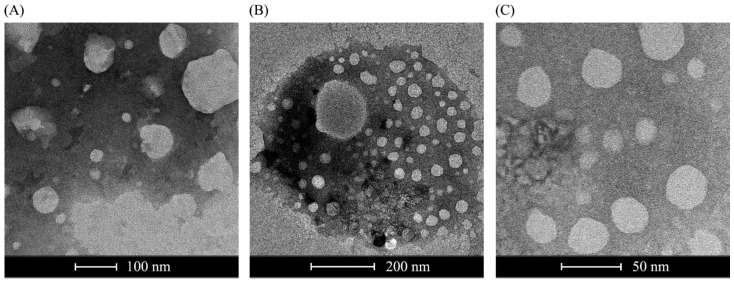
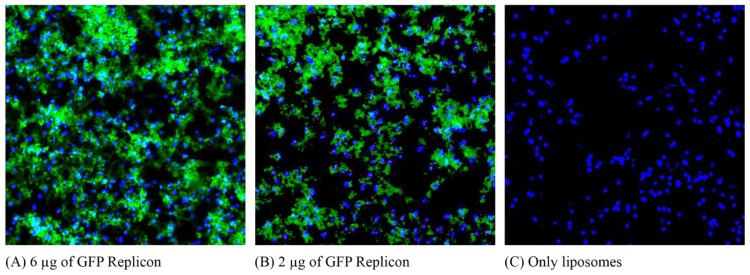
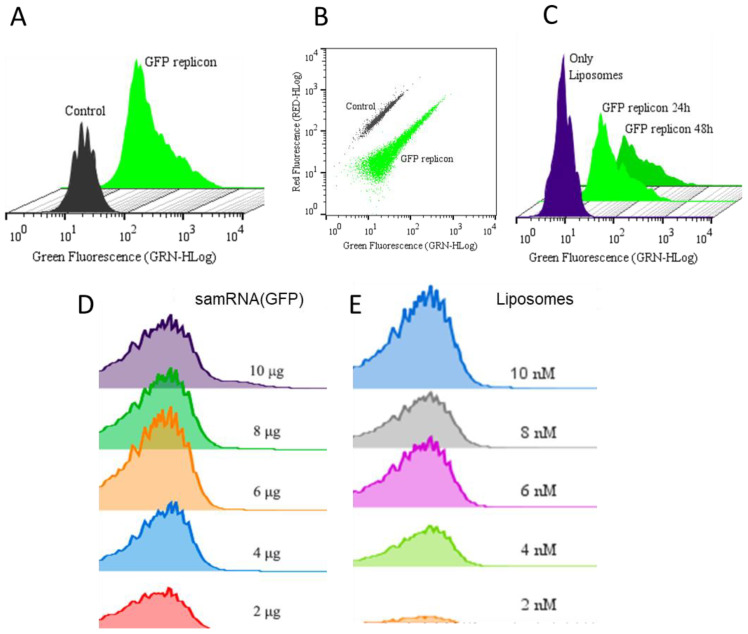
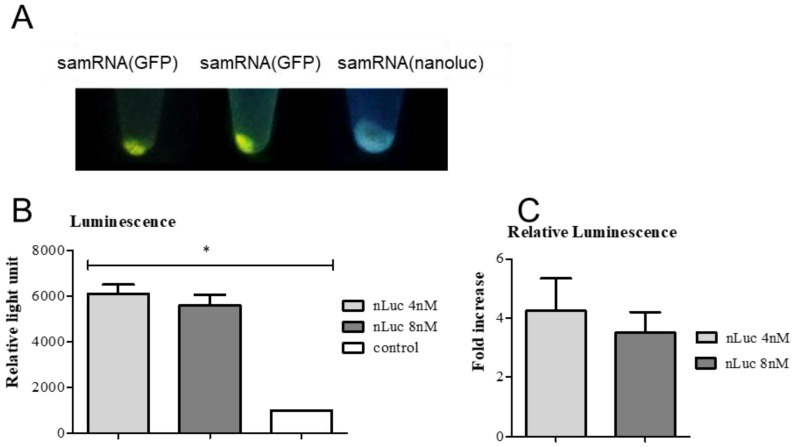
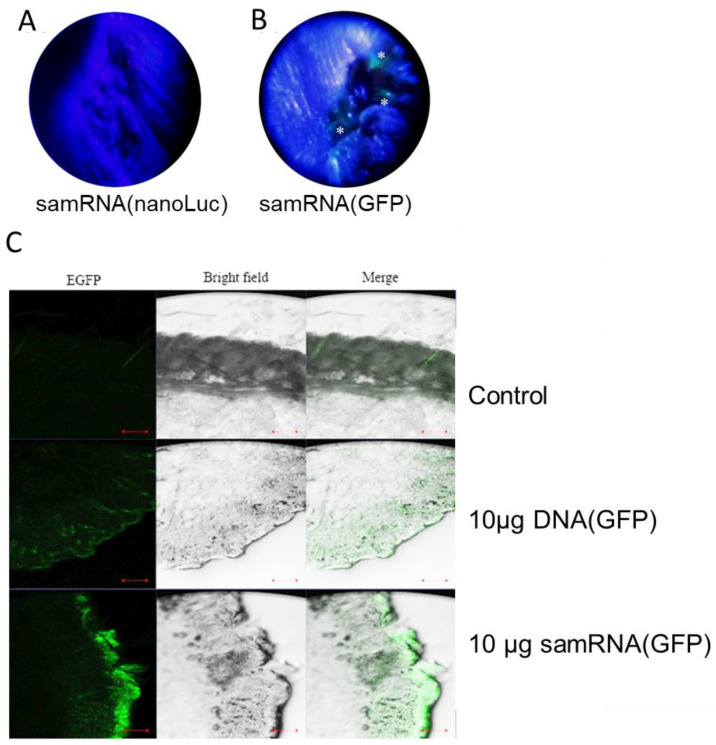
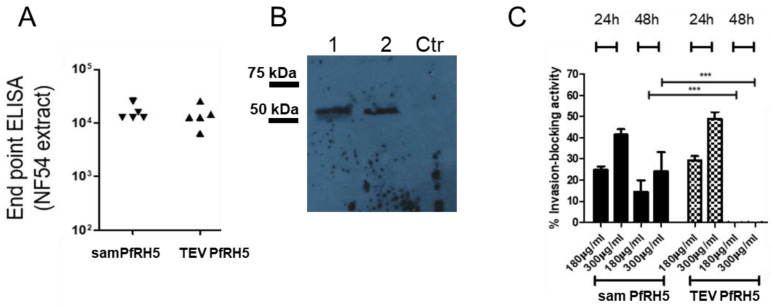
Results: The liposome-replicon complexes showed high transfection efficiencies with in vitro cultured cells, whereas tattooing immunization with GFP-encoding replicons demonstrated gene expression in mouse skin up to 48 h after immunization. Mice immunized with liposomal PfRH5-encoding RNA replicons elicited antibodies that recognized the native protein expressed in P. falciparum schizont extracts, and inhibited the growth of the parasite in vitro.
Conclusion: Intradermal delivery of cationic lipid-encapsulated samRNA constructs is a feasible approach for developing future malaria vaccines.
Keywords: PfRH5; RNA replicons; cationic liposomes; intradermal immunization; malaria; nucleic acid vaccines; tattooing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
