

Stabilization of the Quadruplex-Forming G-Rich Sequences in the Rhinovirus Genome Inhibits Uncoating-Role of Na+ and K
- PMID: 37112983
- PMCID: PMC10141139
- DOI: 10.3390/v15041003
Stabilization of the Quadruplex-Forming G-Rich Sequences in the Rhinovirus Genome Inhibits Uncoating-Role of Na+ and K
Abstract
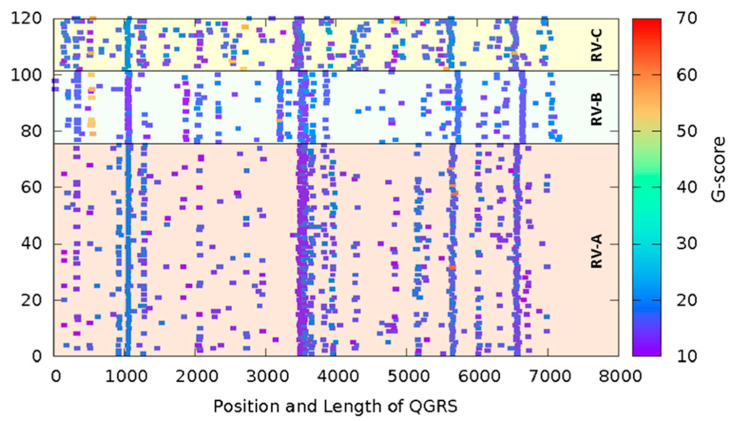
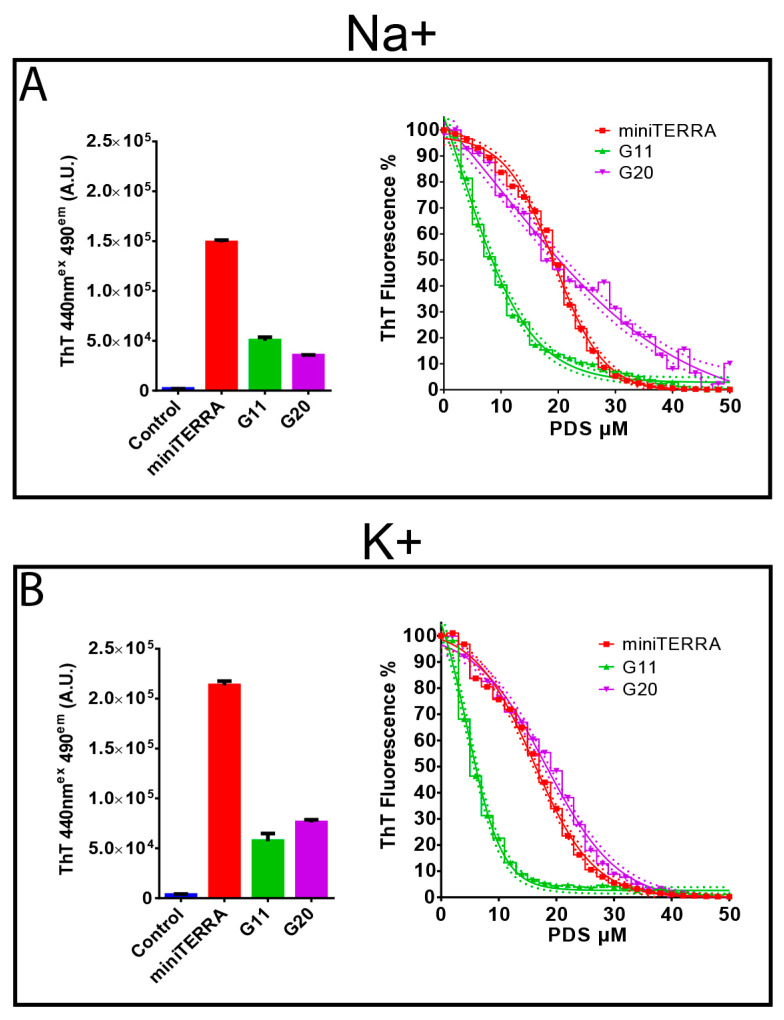
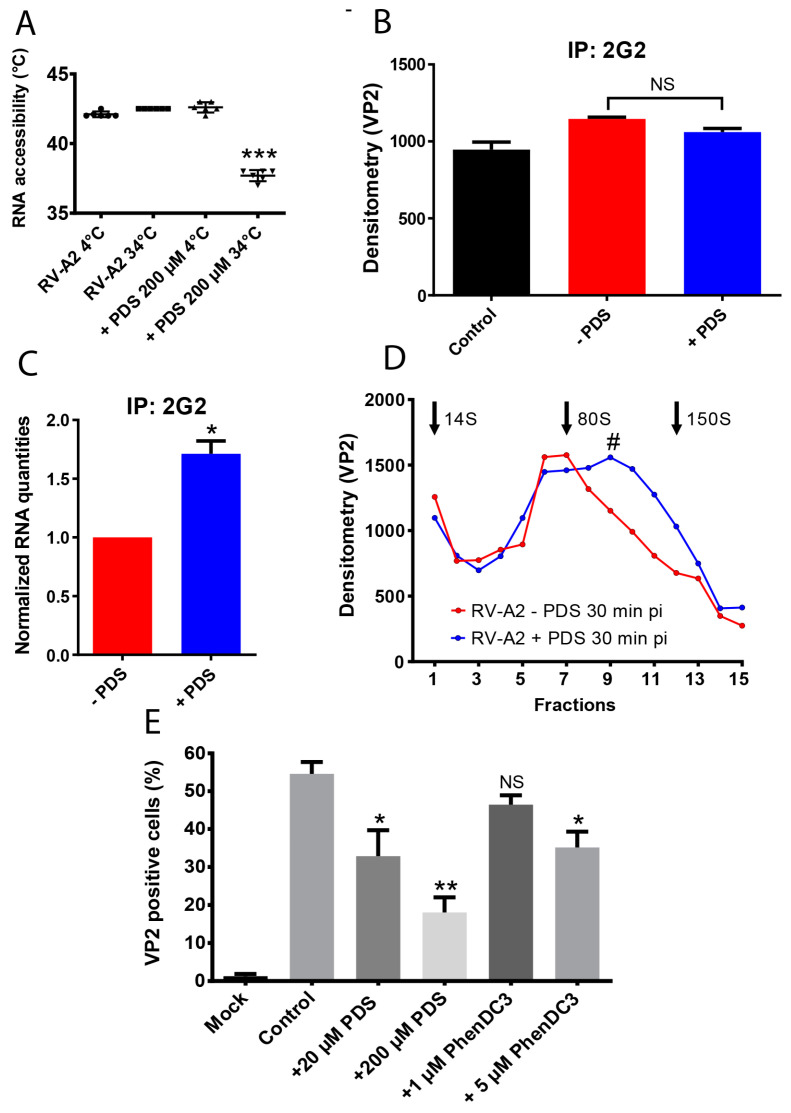
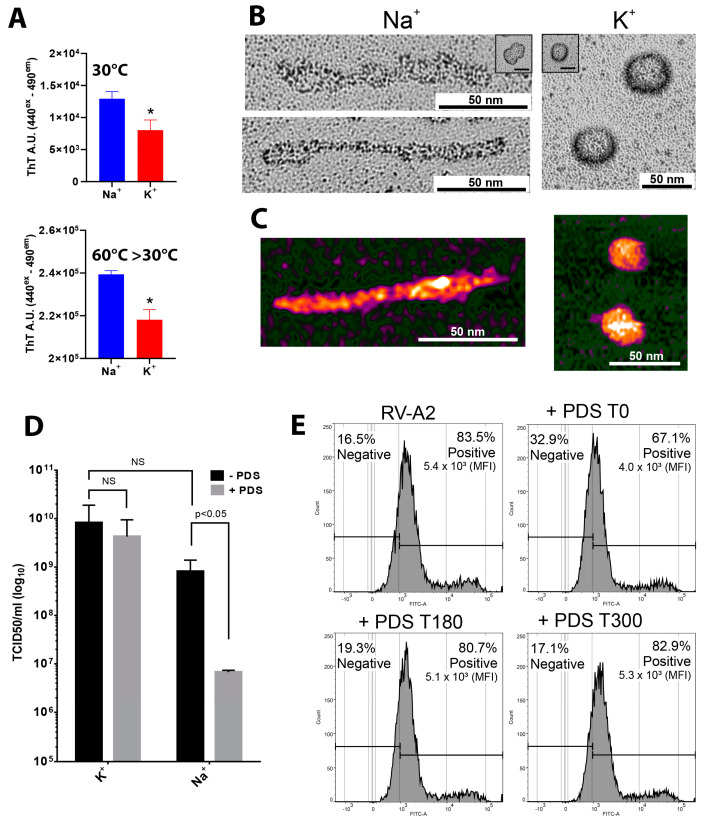
Rhinoviruses (RVs) are the major cause of common cold, a respiratory disease that generally takes a mild course. However, occasionally, RV infection can lead to serious complications in patients debilitated by other ailments, e.g., asthma. Colds are a huge socioeconomic burden as neither vaccines nor other treatments are available. The many existing drug candidates either stabilize the capsid or inhibit the viral RNA polymerase, the viral proteinases, or the functions of other non-structural viral proteins; however, none has been approved by the FDA. Focusing on the genomic RNA as a possible target for antivirals, we asked whether stabilizing RNA secondary structures might inhibit the viral replication cycle. These secondary structures include G-quadruplexes (GQs), which are guanine-rich sequence stretches forming planar guanine tetrads via Hoogsteen base pairing with two or more of them stacking on top of each other; a number of small molecular drug candidates increase the energy required for their unfolding. The propensity of G-quadruplex formation can be predicted with bioinformatics tools and is expressed as a GQ score. Synthetic RNA oligonucleotides derived from the RV-A2 genome with sequences corresponding to the highest and lowest GQ scores indeed exhibited characteristics of GQs. In vivo, the GQ-stabilizing compounds, pyridostatin and PhenDC3, interfered with viral uncoating in Na+ but not in K+-containing phosphate buffers. The thermostability studies and ultrastructural imaging of protein-free viral RNA cores suggest that Na+ keeps the encapsulated genome more open, allowing PDS and PhenDC3 to diffuse into the quasi-crystalline RNA and promote the formation and/or stabilization of GQs; the resulting conformational changes impair RNA unraveling and release from the virion. Preliminary reports have been published.
Keywords: G-quadruplex; K+; Na+; PhenDC3; RNA; folding; inhibition; picornavirus; pyridostatin; rhinovirus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Rhinovirus Inhibitors: Including a New Target, the Viral RNA.Viruses. 2021 Sep 7;13(9):1784. doi: 10.3390/v13091784. Viruses. 2021. PMID: 34578365 Free PMC article. Review.
-
Characterization of G-Quadruplexes in Enterovirus A71 Genome and Their Interaction with G-Quadruplex Ligands.Microbiol Spectr. 2022 Jun 29;10(3):e0046022. doi: 10.1128/spectrum.00460-22. Epub 2022 Apr 21. Microbiol Spectr. 2022. PMID: 35446122 Free PMC article.
-
A RNA Dodecahedral Cage Inside a Human Virus Plays a Dual Biological Role in Virion Assembly and Genome Release Control.J Mol Biol. 2025 Feb 1;437(3):168922. doi: 10.1016/j.jmb.2024.168922. Epub 2024 Dec 24. J Mol Biol. 2025. PMID: 39725271
-
Folding of guanine quadruplex molecules-funnel-like mechanism or kinetic partitioning? An overview from MD simulation studies.Biochim Biophys Acta Gen Subj. 2017 May;1861(5 Pt B):1246-1263. doi: 10.1016/j.bbagen.2016.12.008. Epub 2016 Dec 13. Biochim Biophys Acta Gen Subj. 2017. PMID: 27979677 Review.
-
Same fold, different properties: polarizable molecular dynamics simulations of telomeric and TERRA G-quadruplexes.Nucleic Acids Res. 2020 Jan 24;48(2):561-575. doi: 10.1093/nar/gkz1154. Nucleic Acids Res. 2020. PMID: 31807754 Free PMC article.
Cited by
-
Exosome-mediated regulation of inflammatory pathway during respiratory viral disease.Virol J. 2024 Jan 25;21(1):30. doi: 10.1186/s12985-024-02297-y. Virol J. 2024. PMID: 38273382 Free PMC article. Review.
References
-
- Tang J.W., Lam T.T., Zaraket H., Lipkin W.I., Drews S.J., Hatchette T.F., Heraud J.-M., Koopmans M.P., Abraham A.M., Baraket A., et al. Global epidemiology of non-influenza RNA respiratory viruses: Data gaps and a growing need for surveillance. Lancet Infect. Dis. 2017;17:e320–e326. doi: 10.1016/S1473-3099(17)30238-4. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
