

CD90 is not constitutively expressed in functional innate lymphoid cells
- PMID: 37114052
- PMCID: PMC10126679
- DOI: 10.3389/fimmu.2023.1113735
CD90 is not constitutively expressed in functional innate lymphoid cells
Abstract
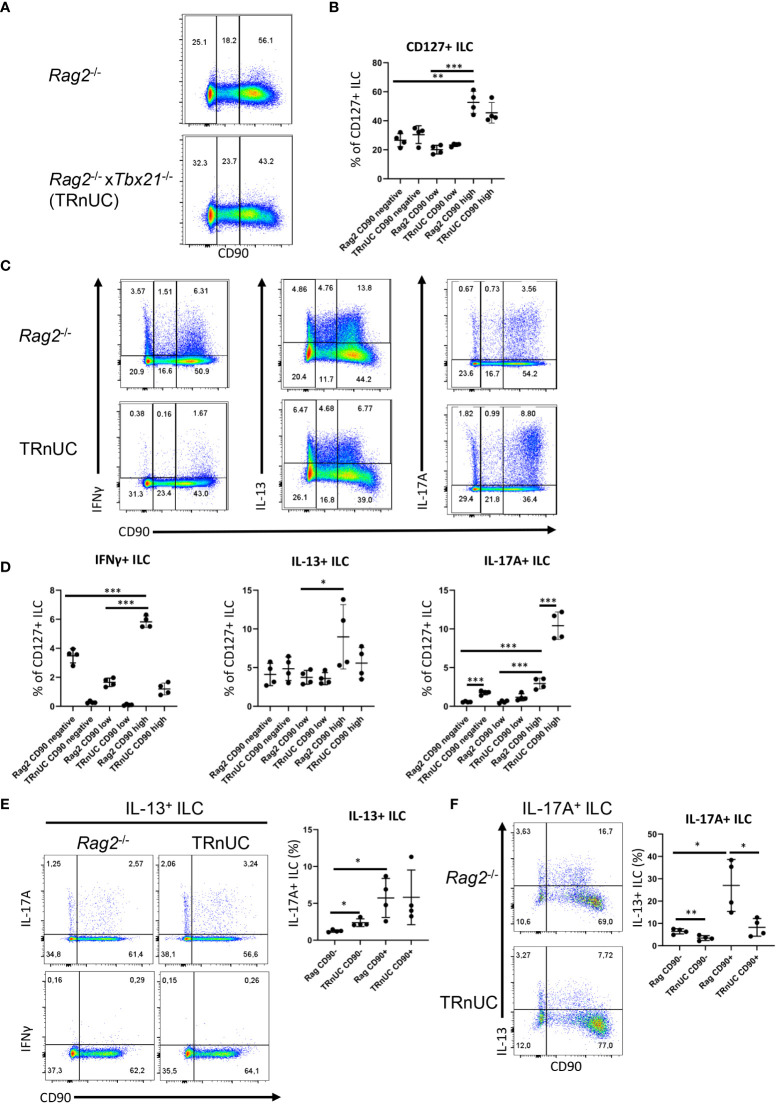
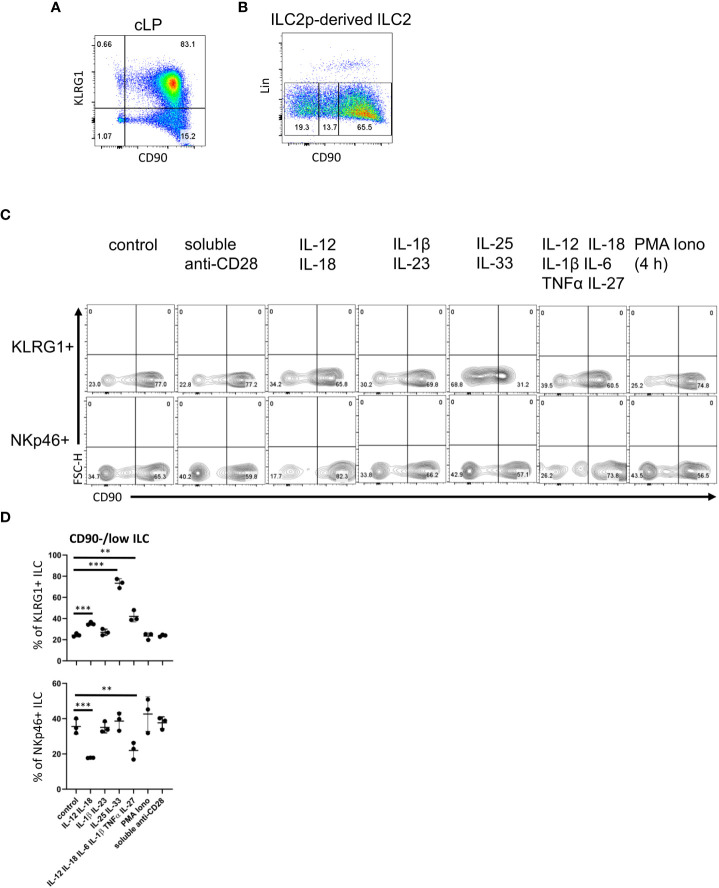
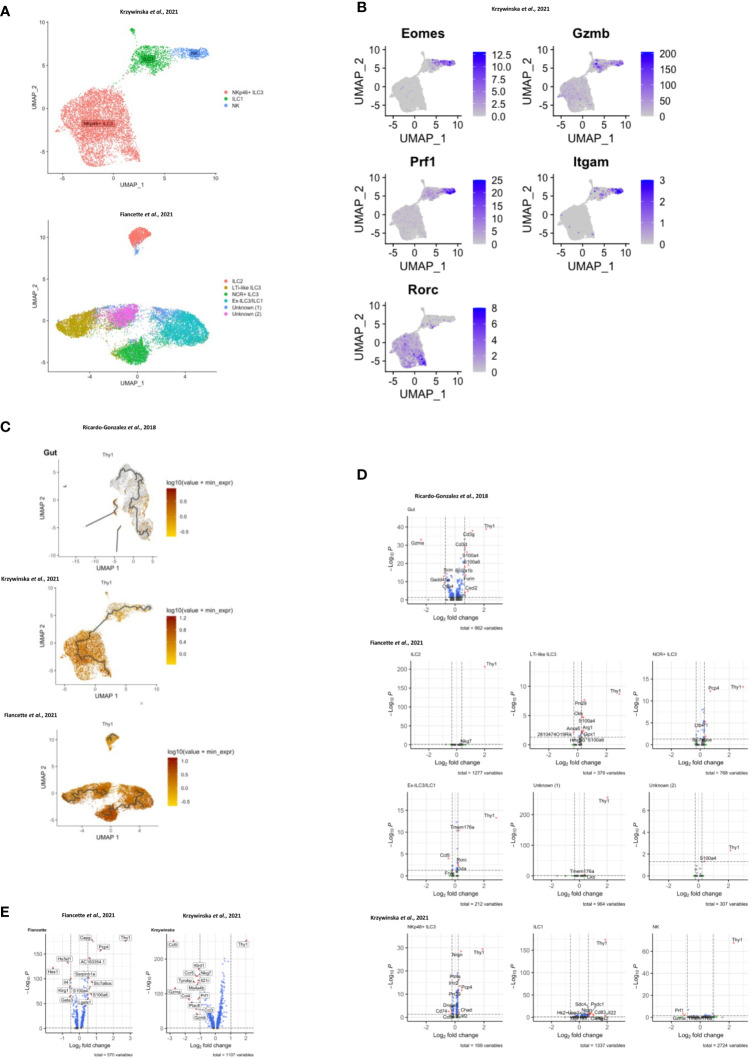
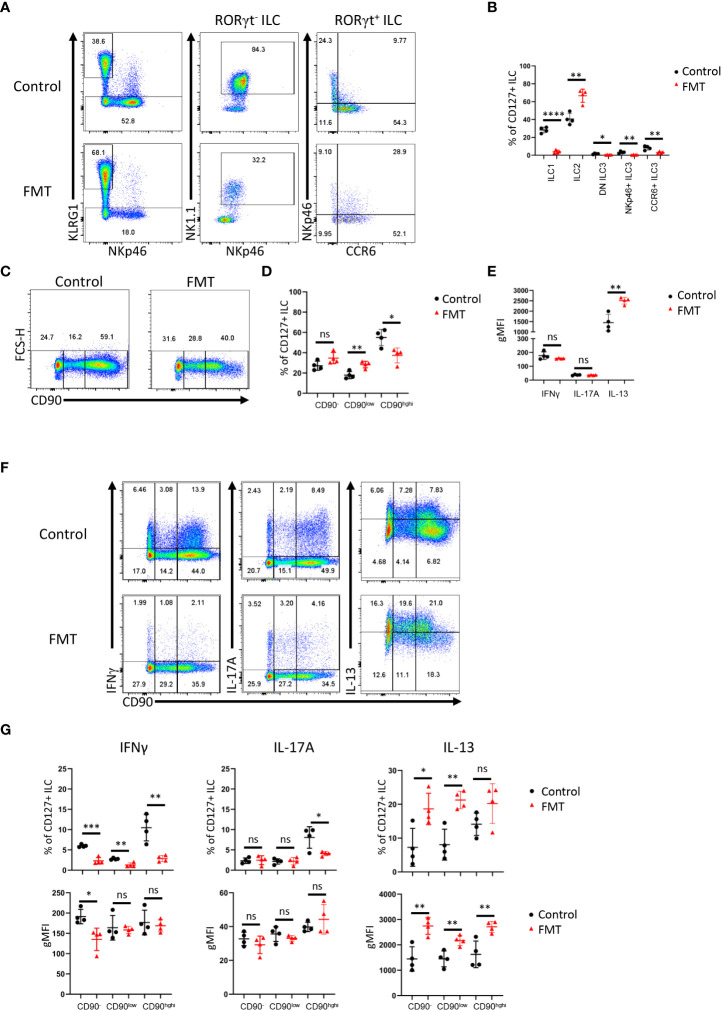
Huge progress has been made in understanding the biology of innate lymphoid cells (ILC) by adopting several well-known concepts in T cell biology. As such, flow cytometry gating strategies and markers, such as CD90, have been applied to indentify ILC. Here, we report that most non-NK intestinal ILC have a high expression of CD90 as expected, but surprisingly a sub-population of cells exhibit low or even no expression of this marker. CD90-negative and CD90-low CD127+ ILC were present amongst all ILC subsets in the gut. The frequency of CD90-negative and CD90-low CD127+ ILC was dependent on stimulatory cues in vitro and enhanced by dysbiosis in vivo. CD90-negative and CD90-low CD127+ ILC were a potential source of IL-13, IFNγ and IL-17A at steady state and upon dysbiosis- and dextran sulphate sodium-elicited colitis. Hence, this study reveals that, contrary to expectations, CD90 is not constitutively expressed by functional ILC in the gut.
Keywords: CD90; DSS-colitis; fecal microbial transplant (FMT); innate lymphoid cell (ILC); intestine.
Copyright © 2023 Schroeder, Beattie, Lo, Zabinski, Powell, Neves, Jenner and Lord.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials